ISSN 0439-755X
CN 11-1911/B
主办:中国心理学会
中国科学院心理研究所
出版:科学出版社
CN 11-1911/B
主办:中国心理学会
中国科学院心理研究所
出版:科学出版社
心理学报 ›› 2023, Vol. 55 ›› Issue (5): 740-751.doi: 10.3724/SP.J.1041.2023.00740 cstr: 32110.14.2023.00740
李彧1,2,3, 位东涛1,2, 邱江1,2( )
)
收稿日期:2022-01-28
发布日期:2023-02-14
出版日期:2023-05-25
通讯作者:
邱江, E-mail: qiuj318@swu.edu.cn基金资助:
LI Yu1,2,3, WEI Dongtao1,2, QIU Jiang1,2()
Received:2022-01-28
Online:2023-02-14
Published:2023-05-25
摘要:
本研究采用功能随机森林的方法, 将聚类过程与抑郁症诊断相结合, 分别在抑郁症和控制组中识别了人格类型(神经质和外向性的组合), 并进一步探究了不同人格类型的静息态功能连接差异。聚类分析结果显示, 抑郁症以高神经质和低外向性趋势的个体为主, 但同样有低神经质和高外向性趋势的个体。控制组样本则以低神经质和高外向性个体为主。静息态功能连接的结果显示:在不考虑人格亚型的情况下, 抑郁症和控制组在杏仁核/海马/脑岛−边缘网络/默认网络/控制网络的功能连接上均无显著差异。在纳入聚类分析所划分的亚型进行统计后, 多种人格类型在左侧杏仁核/脑岛−边缘网络(以眶额皮质区域为主)的功能连接强度上呈现出显著差异。本研究基于个人视角识别的抑郁症人格类型更符合现实情况与个体认知模式, 具有潜在的临床应用价值, 并且其功能连接的差异对理解抑郁症异质性提供了神经层面的参考。
中图分类号:
李彧, 位东涛, 邱江. (2023). 抑郁症的人格类型及其脑功能连接基础. 心理学报, 55(5), 740-751.
LI Yu, WEI Dongtao, QIU Jiang. (2023). Personality subtypes of depressive disorders and their functional connectivity basis. Acta Psychologica Sinica, 55(5), 740-751.
| 组别 | 年龄 | 性别 | 教育年限 | HAMD | |||||
|---|---|---|---|---|---|---|---|---|---|
| 均值 | 标准差 | 范围 | 男 | 女 | 均值 | 标准差 | 均值 | 标准差 | |
| 抑郁 症 | 38.9 | 13.3 | 18~71 | 61 | 98 | 11.7 | 3.69 | 21.1 | 5.02 |
| 控制 组 | 41.7 | 15.9 | 19~70 | 54 | 102 | 12.3 | 4.53 | 2.17 | 1.87 |
| 统计 值 | t(313) = 1.68 p = 0.094 | χ²(1) = 0.478 p = 0.490 | t(313) = 1.23 p = 0.22 | t(313) = 44.08 p < 0.001 | |||||
表1 基本人口学信息
| 组别 | 年龄 | 性别 | 教育年限 | HAMD | |||||
|---|---|---|---|---|---|---|---|---|---|
| 均值 | 标准差 | 范围 | 男 | 女 | 均值 | 标准差 | 均值 | 标准差 | |
| 抑郁 症 | 38.9 | 13.3 | 18~71 | 61 | 98 | 11.7 | 3.69 | 21.1 | 5.02 |
| 控制 组 | 41.7 | 15.9 | 19~70 | 54 | 102 | 12.3 | 4.53 | 2.17 | 1.87 |
| 统计 值 | t(313) = 1.68 p = 0.094 | χ²(1) = 0.478 p = 0.490 | t(313) = 1.23 p = 0.22 | t(313) = 44.08 p < 0.001 | |||||
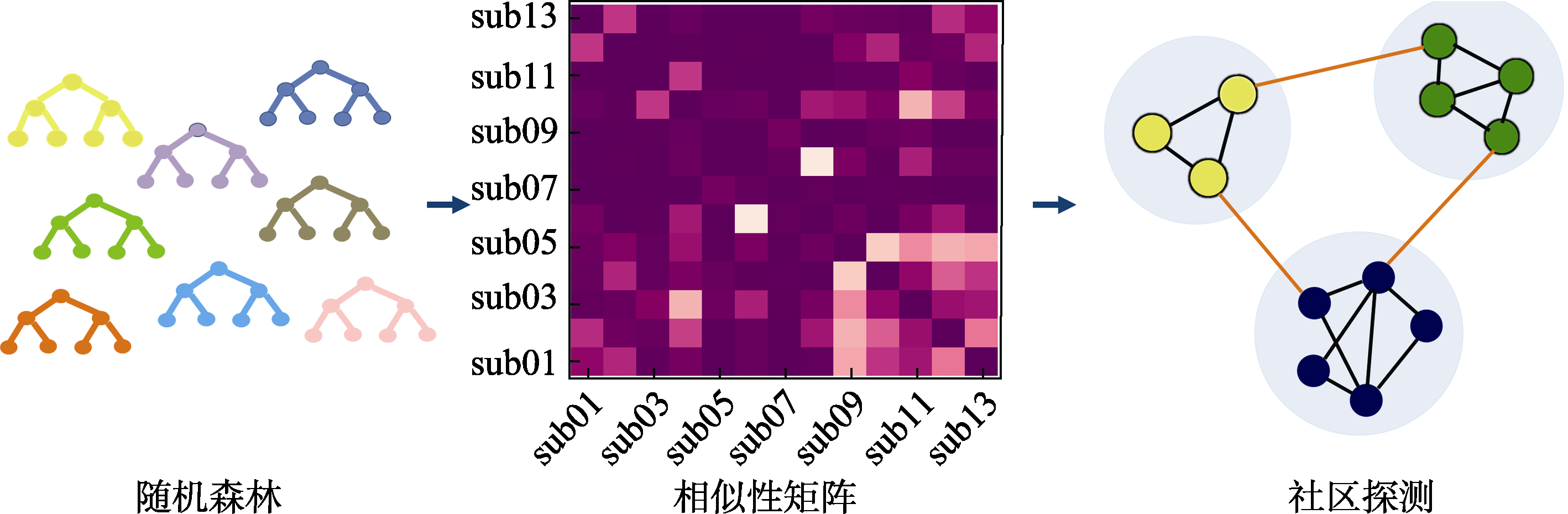
图1 聚类流程 注:随机森林:单颗决策树上的内部节点代表自变量(即输入特征), 节点之间的连接代表决策, 终端节点(叶节点)代表一个结果(即预测变量)。相似性矩阵:行和列代表被试, 每个单元格代表在所有树中成对被试落到相同终端节点的概率。
| 组间比较 | 神经质 | 外向性 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| t | pholm | Cohen's d | 95% CI | t | pholm | Cohen's d | 95% CI | |||||
| DD1 | − | DD2 | −13.91 | <0.001 | −3.087 | −3.588 | −2.585 | 12.85 | <0.001 | 2.851 | 2.359 | 3.344 |
| − | DD3 | −13.76 | <0.001 | −3.076 | −3.579 | −2.572 | −0.03 | 1.000 | −0.006 | −0.446 | 0.433 | |
| − | DD4 | −15.45 | <0.001 | −3.654 | −4.204 | −3.105 | 6.25 | <0.001 | 1.478 | 0.998 | 1.958 | |
| − | CON1 | 5.78 | <0.001 | 1.247 | 0.811 | 1.683 | −4.44 | <0.001 | −0.958 | −1.389 | −0.527 | |
| − | CON2 | −3.07 | 0.024 | −0.729 | −1.200 | −0.257 | 2.63 | 0.124 | 0.626 | 0.156 | 1.097 | |
| − | CON3 | 3.92 | 0.001 | 0.946 | 0.466 | 1.426 | 0.58 | 1.000 | 0.139 | −0.335 | 0.614 | |
| − | CON4 | −2.49 | 0.106 | −0.756 | −1.355 | −0.156 | −2.26 | 0.267 | −0.687 | −1.286 | −0.088 | |
| − | CON5 | −7.69 | <0.001 | −2.729 | −3.460 | −1.998 | 0.88 | 1.000 | 0.313 | −0.385 | 1.012 | |
| DD2 | − | DD3 | 0.05 | 1.000 | 0.011 | −0.415 | 0.437 | −13.20 | <0.001 | −2.858 | −3.341 | −2.374 |
| − | DD4 | −2.41 | 0.110 | −0.567 | −1.033 | −0.102 | −5.83 | <0.001 | −1.373 | −1.849 | −0.897 | |
| − | CON1 | 19.44 | <0.001 | 4.334 | 3.775 | 4.893 | −17.08 | <0.001 | −3.809 | −4.343 | −3.275 | |
| − | CON2 | 9.29 | <0.001 | 2.358 | 1.824 | 2.892 | −8.76 | <0.001 | −2.225 | −2.755 | −1.695 | |
| − | CON3 | 18.15 | <0.001 | 4.033 | 3.490 | 4.576 | −12.21 | <0.001 | −2.712 | −3.200 | −2.224 | |
| − | CON4 | 7.77 | <0.001 | 2.331 | 1.713 | 2.950 | −11.80 | <0.001 | −3.538 | −4.192 | −2.884 | |
| − | CON5 | 1.04 | 1.000 | 0.358 | −0.318 | 1.034 | −7.39 | <0.001 | −2.538 | −3.243 | −1.833 | |
| DD3 | − | DD4 | −2.43 | 0.110 | −0.579 | −1.049 | −0.108 | 6.24 | <0.001 | 1.485 | 1.001 | 1.968 |
| − | CON1 | 19.36 | <0.001 | 4.323 | 3.764 | 4.882 | −4.26 | <0.001 | −0.952 | −1.397 | −0.506 | |
| − | CON2 | 9.29 | <0.001 | 2.347 | 1.816 | 2.878 | 2.51 | 0.166 | 0.633 | 0.133 | 1.132 | |
| − | CON3 | 17.56 | <0.001 | 4.022 | 3.468 | 4.575 | 0.64 | 1.000 | 0.146 | −0.305 | 0.597 | |
| − | CON4 | 7.68 | <0.001 | 2.320 | 1.698 | 2.943 | −2.25 | 0.267 | −0.680 | −1.277 | −0.083 | |
| − | CON5 | 0.99 | 1.000 | 0.347 | −0.340 | 1.033 | 0.92 | 1.000 | 0.320 | −0.367 | 1.006 | |
| DD4 | − | CON1 | 21.03 | <0.001 | 4.901 | 4.298 | 5.505 | −10.45 | <0.001 | −2.436 | −2.934 | −1.938 |
| − | CON2 | 11.43 | <0.001 | 2.925 | 2.370 | 3.481 | −3.33 | 0.017 | −0.852 | −1.360 | −0.344 | |
| − | CON3 | 18.30 | <0.001 | 4.600 | 3.984 | 5.217 | −5.33 | <0.001 | −1.339 | −1.845 | −0.833 | |
| − | CON4 | 9.23 | <0.001 | 2.899 | 2.239 | 3.559 | −6.89 | <0.001 | −2.165 | −2.807 | −1.523 | |
| − | CON5 | 2.56 | 0.098 | 0.925 | 0.210 | 1.640 | −3.22 | 0.022 | −1.165 | −1.882 | −0.448 | |
| CON1 | − | CON2 | −8.88 | <0.001 | −1.976 | −2.442 | −1.510 | 7.12 | <0.001 | 1.584 | 1.128 | 2.040 |
| − | CON3 | −1.21 | 1.000 | −0.301 | −0.791 | 0.189 | 4.41 | <0.001 | 1.097 | 0.600 | 1.595 | |
| − | CON4 | −6.63 | <0.001 | −2.003 | −2.618 | −1.387 | 0.90 | 1.000 | 0.271 | −0.324 | 0.866 | |
| − | CON5 | −11.06 | <0.001 | −3.976 | −4.752 | −3.201 | 3.54 | 0.008 | 1.271 | 0.556 | 1.986 | |
| CON2 | − | CON3 | 5.85 | <0.001 | 1.675 | 1.096 | 2.254 | −1.70 | 0.810 | −0.487 | −1.052 | 0.078 |
| − | CON4 | −0.08 | 1.000 | −0.027 | −0.661 | 0.608 | −4.07 | 0.001 | −1.313 | −1.956 | −0.670 | |
| − | CON5 | −5.19 | <0.001 | −2.000 | −2.775 | −1.225 | −0.81 | 1.000 | −0.313 | −1.072 | 0.446 | |
| CON3 | − | CON4 | −5.51 | <0.001 | −1.702 | −2.324 | −1.079 | −2.68 | 0.118 | −0.826 | −1.437 | −0.215 |
| − | CON5 | −10.88 | <0.001 | −3.675 | −4.402 | −2.948 | 0.52 | 1.000 | 0.174 | −0.491 | 0.839 | |
| CON4 | − | CON5 | −4.88 | <0.001 | −1.974 | −2.784 | −1.163 | 2.47 | 0.167 | 1.000 | 0.201 | 1.799 |
表S1 各人格类型在神经质和外向性上的组间差异
| 组间比较 | 神经质 | 外向性 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| t | pholm | Cohen's d | 95% CI | t | pholm | Cohen's d | 95% CI | |||||
| DD1 | − | DD2 | −13.91 | <0.001 | −3.087 | −3.588 | −2.585 | 12.85 | <0.001 | 2.851 | 2.359 | 3.344 |
| − | DD3 | −13.76 | <0.001 | −3.076 | −3.579 | −2.572 | −0.03 | 1.000 | −0.006 | −0.446 | 0.433 | |
| − | DD4 | −15.45 | <0.001 | −3.654 | −4.204 | −3.105 | 6.25 | <0.001 | 1.478 | 0.998 | 1.958 | |
| − | CON1 | 5.78 | <0.001 | 1.247 | 0.811 | 1.683 | −4.44 | <0.001 | −0.958 | −1.389 | −0.527 | |
| − | CON2 | −3.07 | 0.024 | −0.729 | −1.200 | −0.257 | 2.63 | 0.124 | 0.626 | 0.156 | 1.097 | |
| − | CON3 | 3.92 | 0.001 | 0.946 | 0.466 | 1.426 | 0.58 | 1.000 | 0.139 | −0.335 | 0.614 | |
| − | CON4 | −2.49 | 0.106 | −0.756 | −1.355 | −0.156 | −2.26 | 0.267 | −0.687 | −1.286 | −0.088 | |
| − | CON5 | −7.69 | <0.001 | −2.729 | −3.460 | −1.998 | 0.88 | 1.000 | 0.313 | −0.385 | 1.012 | |
| DD2 | − | DD3 | 0.05 | 1.000 | 0.011 | −0.415 | 0.437 | −13.20 | <0.001 | −2.858 | −3.341 | −2.374 |
| − | DD4 | −2.41 | 0.110 | −0.567 | −1.033 | −0.102 | −5.83 | <0.001 | −1.373 | −1.849 | −0.897 | |
| − | CON1 | 19.44 | <0.001 | 4.334 | 3.775 | 4.893 | −17.08 | <0.001 | −3.809 | −4.343 | −3.275 | |
| − | CON2 | 9.29 | <0.001 | 2.358 | 1.824 | 2.892 | −8.76 | <0.001 | −2.225 | −2.755 | −1.695 | |
| − | CON3 | 18.15 | <0.001 | 4.033 | 3.490 | 4.576 | −12.21 | <0.001 | −2.712 | −3.200 | −2.224 | |
| − | CON4 | 7.77 | <0.001 | 2.331 | 1.713 | 2.950 | −11.80 | <0.001 | −3.538 | −4.192 | −2.884 | |
| − | CON5 | 1.04 | 1.000 | 0.358 | −0.318 | 1.034 | −7.39 | <0.001 | −2.538 | −3.243 | −1.833 | |
| DD3 | − | DD4 | −2.43 | 0.110 | −0.579 | −1.049 | −0.108 | 6.24 | <0.001 | 1.485 | 1.001 | 1.968 |
| − | CON1 | 19.36 | <0.001 | 4.323 | 3.764 | 4.882 | −4.26 | <0.001 | −0.952 | −1.397 | −0.506 | |
| − | CON2 | 9.29 | <0.001 | 2.347 | 1.816 | 2.878 | 2.51 | 0.166 | 0.633 | 0.133 | 1.132 | |
| − | CON3 | 17.56 | <0.001 | 4.022 | 3.468 | 4.575 | 0.64 | 1.000 | 0.146 | −0.305 | 0.597 | |
| − | CON4 | 7.68 | <0.001 | 2.320 | 1.698 | 2.943 | −2.25 | 0.267 | −0.680 | −1.277 | −0.083 | |
| − | CON5 | 0.99 | 1.000 | 0.347 | −0.340 | 1.033 | 0.92 | 1.000 | 0.320 | −0.367 | 1.006 | |
| DD4 | − | CON1 | 21.03 | <0.001 | 4.901 | 4.298 | 5.505 | −10.45 | <0.001 | −2.436 | −2.934 | −1.938 |
| − | CON2 | 11.43 | <0.001 | 2.925 | 2.370 | 3.481 | −3.33 | 0.017 | −0.852 | −1.360 | −0.344 | |
| − | CON3 | 18.30 | <0.001 | 4.600 | 3.984 | 5.217 | −5.33 | <0.001 | −1.339 | −1.845 | −0.833 | |
| − | CON4 | 9.23 | <0.001 | 2.899 | 2.239 | 3.559 | −6.89 | <0.001 | −2.165 | −2.807 | −1.523 | |
| − | CON5 | 2.56 | 0.098 | 0.925 | 0.210 | 1.640 | −3.22 | 0.022 | −1.165 | −1.882 | −0.448 | |
| CON1 | − | CON2 | −8.88 | <0.001 | −1.976 | −2.442 | −1.510 | 7.12 | <0.001 | 1.584 | 1.128 | 2.040 |
| − | CON3 | −1.21 | 1.000 | −0.301 | −0.791 | 0.189 | 4.41 | <0.001 | 1.097 | 0.600 | 1.595 | |
| − | CON4 | −6.63 | <0.001 | −2.003 | −2.618 | −1.387 | 0.90 | 1.000 | 0.271 | −0.324 | 0.866 | |
| − | CON5 | −11.06 | <0.001 | −3.976 | −4.752 | −3.201 | 3.54 | 0.008 | 1.271 | 0.556 | 1.986 | |
| CON2 | − | CON3 | 5.85 | <0.001 | 1.675 | 1.096 | 2.254 | −1.70 | 0.810 | −0.487 | −1.052 | 0.078 |
| − | CON4 | −0.08 | 1.000 | −0.027 | −0.661 | 0.608 | −4.07 | 0.001 | −1.313 | −1.956 | −0.670 | |
| − | CON5 | −5.19 | <0.001 | −2.000 | −2.775 | −1.225 | −0.81 | 1.000 | −0.313 | −1.072 | 0.446 | |
| CON3 | − | CON4 | −5.51 | <0.001 | −1.702 | −2.324 | −1.079 | −2.68 | 0.118 | −0.826 | −1.437 | −0.215 |
| − | CON5 | −10.88 | <0.001 | −3.675 | −4.402 | −2.948 | 0.52 | 1.000 | 0.174 | −0.491 | 0.839 | |
| CON4 | − | CON5 | −4.88 | <0.001 | −1.974 | −2.784 | −1.163 | 2.47 | 0.167 | 1.000 | 0.201 | 1.799 |
| 左脑岛- LimbicB | 左脑岛- DMNA | 左脑岛- DMNB | 右脑岛- LimbicB | 右脑岛- DMNA | 右脑岛- DMNB | 外向性 | ||
|---|---|---|---|---|---|---|---|---|
| 左脑岛-LimbicB | 皮尔逊相关系数 | — | ||||||
| p | — | |||||||
| 95% CI | — | |||||||
| — | ||||||||
| 左脑岛-DMNA | 皮尔逊相关系数 | 0.689 | — | |||||
| p | <0.001 | — | ||||||
| 95% CI | 0.749 | — | ||||||
| 0.617 | — | |||||||
| 左脑岛-DMNB | 皮尔逊相关系数 | 0.688 | 0.863 | — | ||||
| p | <0.001 | <0.001 | — | |||||
| 95% CI | 0.748 | 0.892 | — | |||||
| 0.616 | 0.828 | — | ||||||
| 右脑岛-LimbicB | 皮尔逊相关系数 | 0.902 | 0.617 | 0.614 | — | |||
| p | <0.001 | <0.001 | <0.001 | — | ||||
| 95% CI | 0.923 | 0.689 | 0.686 | — | ||||
| 0.875 | 0.533 | 0.53 | — | |||||
| 右脑岛-DMNA | 皮尔逊相关系数 | 0.649 | 0.913 | 0.795 | 0.698 | — | ||
| p | <0.001 | <0.001 | <0.001 | <0.001 | — | |||
| 95% CI | 0.716 | 0.932 | 0.837 | 0.757 | — | |||
| 0.571 | 0.89 | 0.744 | 0.627 | — | ||||
| 右脑岛-DMNB | 皮尔逊相关系数 | 0.658 | 0.809 | 0.921 | 0.686 | 0.862 | — | |
| p | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | — | ||
| 95% CI | 0.724 | 0.848 | 0.938 | 0.747 | 0.891 | — | ||
| 0.581 | 0.761 | 0.9 | 0.614 | 0.826 | — | |||
| 外向性 | 皮尔逊相关系数 | 0.204 | 0.208 | 0.208 | 0.214 | 0.182 | 0.223 | — |
| p | 0.001 | 0.001 | <0.001 | <0.001 | 0.004 | <0.001 | — | |
| 95% CI | 0.32 | 0.324 | 0.325 | 0.33 | 0.3 | 0.339 | — | |
| 0.081 | 0.085 | 0.086 | 0.092 | 0.059 | 0.101 | — |
表S2 外向性与功能连接的相关矩阵
| 左脑岛- LimbicB | 左脑岛- DMNA | 左脑岛- DMNB | 右脑岛- LimbicB | 右脑岛- DMNA | 右脑岛- DMNB | 外向性 | ||
|---|---|---|---|---|---|---|---|---|
| 左脑岛-LimbicB | 皮尔逊相关系数 | — | ||||||
| p | — | |||||||
| 95% CI | — | |||||||
| — | ||||||||
| 左脑岛-DMNA | 皮尔逊相关系数 | 0.689 | — | |||||
| p | <0.001 | — | ||||||
| 95% CI | 0.749 | — | ||||||
| 0.617 | — | |||||||
| 左脑岛-DMNB | 皮尔逊相关系数 | 0.688 | 0.863 | — | ||||
| p | <0.001 | <0.001 | — | |||||
| 95% CI | 0.748 | 0.892 | — | |||||
| 0.616 | 0.828 | — | ||||||
| 右脑岛-LimbicB | 皮尔逊相关系数 | 0.902 | 0.617 | 0.614 | — | |||
| p | <0.001 | <0.001 | <0.001 | — | ||||
| 95% CI | 0.923 | 0.689 | 0.686 | — | ||||
| 0.875 | 0.533 | 0.53 | — | |||||
| 右脑岛-DMNA | 皮尔逊相关系数 | 0.649 | 0.913 | 0.795 | 0.698 | — | ||
| p | <0.001 | <0.001 | <0.001 | <0.001 | — | |||
| 95% CI | 0.716 | 0.932 | 0.837 | 0.757 | — | |||
| 0.571 | 0.89 | 0.744 | 0.627 | — | ||||
| 右脑岛-DMNB | 皮尔逊相关系数 | 0.658 | 0.809 | 0.921 | 0.686 | 0.862 | — | |
| p | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | — | ||
| 95% CI | 0.724 | 0.848 | 0.938 | 0.747 | 0.891 | — | ||
| 0.581 | 0.761 | 0.9 | 0.614 | 0.826 | — | |||
| 外向性 | 皮尔逊相关系数 | 0.204 | 0.208 | 0.208 | 0.214 | 0.182 | 0.223 | — |
| p | 0.001 | 0.001 | <0.001 | <0.001 | 0.004 | <0.001 | — | |
| 95% CI | 0.32 | 0.324 | 0.325 | 0.33 | 0.3 | 0.339 | — | |
| 0.081 | 0.085 | 0.086 | 0.092 | 0.059 | 0.101 | — |
| 脑模板中的节点编号 | 节点名称 | 坐标 | ||
|---|---|---|---|---|
| R | A | S | ||
| 109 | 17Networks_LH_LimbicB_OFC_1 | -12 | 25 | -21 |
| 110 | 17Networks_LH_LimbicB_OFC_2 | -24 | 23 | -20 |
| 111 | 17Networks_LH_LimbicB_OFC_3 | -10 | 47 | -21 |
| 112 | 17Networks_LH_LimbicB_OFC_4 | -4 | 23 | -19 |
| 113 | 17Networks_LH_LimbicB_OFC_5 | -15 | 65 | -8 |
| 114 | 17Networks_LH_LimbicA_TempPole_1 | -37 | -5 | -42 |
| 115 | 17Networks_LH_LimbicA_TempPole_2 | -25 | 6 | -39 |
| 116 | 17Networks_LH_LimbicA_TempPole_3 | -26 | -9 | -33 |
| 117 | 17Networks_LH_LimbicA_TempPole_4 | -54 | -21 | -31 |
| 118 | 17Networks_LH_LimbicA_TempPole_5 | -40 | -21 | -27 |
| 119 | 17Networks_LH_LimbicA_TempPole_6 | -32 | 12 | -29 |
| 120 | 17Networks_LH_LimbicA_TempPole_7 | -44 | 5 | -17 |
| 121 | 17Networks_LH_ContA_Temp_1 | -55 | -62 | -1 |
| 122 | 17Networks_LH_ContA_IPS_1 | -29 | -74 | 42 |
| 123 | 17Networks_LH_ContA_IPS_2 | -58 | -42 | 45 |
| 124 | 17Networks_LH_ContA_IPS_3 | -35 | -62 | 48 |
| 125 | 17Networks_LH_ContA_IPS_4 | -45 | -41 | 47 |
| 126 | 17Networks_LH_ContA_IPS_5 | -33 | -46 | 41 |
| 127 | 17Networks_LH_ContA_PFCd_1 | -21 | 5 | 65 |
| 128 | 17Networks_LH_ContA_PFClv_1 | -48 | 35 | 10 |
| 129 | 17Networks_LH_ContA_PFClv_2 | -42 | 38 | 22 |
| 130 | 17Networks_LH_ContA_PFCl_1 | -49 | 6 | 26 |
| 131 | 17Networks_LH_ContA_PFCl_2 | -45 | 20 | 27 |
| 132 | 17Networks_LH_ContA_PFCl_3 | -39 | 7 | 34 |
| 133 | 17Networks_LH_ContA_Cingm_1 | -3 | 5 | 29 |
| 134 | 17Networks_LH_ContB_Temp_1 | -60 | -36 | -18 |
| 135 | 17Networks_LH_ContB_Temp_2 | -60 | -49 | -10 |
| 136 | 17Networks_LH_ContB_IPL_1 | -49 | -60 | 47 |
| 137 | 17Networks_LH_ContB_IPL_2 | -53 | -50 | 45 |
| 138 | 17Networks_LH_ContB_IPL_3 | -42 | -52 | 49 |
| 139 | 17Networks_LH_ContB_PFCd_1 | -30 | 14 | 57 |
| 140 | 17Networks_LH_ContB_PFClv_1 | -42 | 49 | -6 |
| 141 | 17Networks_LH_ContB_PFClv_2 | -28 | 58 | -1 |
| 142 | 17Networks_LH_ContB_PFClv_3 | -28 | 57 | 13 |
| 143 | 17Networks_LH_ContB_PFCmp_1 | -4 | 28 | 47 |
| 144 | 17Networks_LH_ContC_pCun_1 | -10 | -70 | 32 |
| 145 | 17Networks_LH_ContC_pCun_2 | -9 | -77 | 45 |
| 146 | 17Networks_LH_ContC_pCun_3 | -5 | -64 | 52 |
| 147 | 17Networks_LH_ContC_Cingp_1 | -6 | -41 | 24 |
| 148 | 17Networks_LH_ContC_Cingp_2 | -4 | -22 | 29 |
| 149 | 17Networks_LH_DefaultA_IPL_1 | -47 | -64 | 31 |
| 150 | 17Networks_LH_DefaultA_IPL_2 | -41 | -72 | 43 |
| 151 | 17Networks_LH_DefaultA_PFCd_1 | -25 | 28 | 43 |
| 152 | 17Networks_LH_DefaultA_PFCd_2 | -18 | 36 | 48 |
| 153 | 17Networks_LH_DefaultA_PFCd_3 | -22 | 20 | 52 |
| 154 | 17Networks_LH_DefaultA_pCunPCC_1 | -4 | -53 | 20 |
| 155 | 17Networks_LH_DefaultA_pCunPCC_2 | -5 | -60 | 30 |
| 156 | 17Networks_LH_DefaultA_pCunPCC_3 | -7 | -44 | 32 |
| 157 | 17Networks_LH_DefaultA_pCunPCC_4 | -4 | -34 | 38 |
| 158 | 17Networks_LH_DefaultA_pCunPCC_5 | -3 | -15 | 37 |
| 159 | 17Networks_LH_DefaultA_pCunPCC_6 | -3 | -68 | 41 |
| 160 | 17Networks_LH_DefaultA_pCunPCC_7 | -7 | -51 | 43 |
| 161 | 17Networks_LH_DefaultA_PFCm_1 | -5 | 55 | -10 |
| 162 | 17Networks_LH_DefaultA_PFCm_2 | -6 | 35 | -9 |
| 163 | 17Networks_LH_DefaultA_PFCm_3 | -6 | 59 | 7 |
| 164 | 17Networks_LH_DefaultA_PFCm_4 | -6 | 45 | 6 |
| 165 | 17Networks_LH_DefaultA_PFCm_5 | -16 | 67 | 8 |
| 166 | 17Networks_LH_DefaultA_PFCm_6 | -5 | 34 | 21 |
| 167 | 17Networks_LH_DefaultB_Temp_1 | -44 | 13 | -34 |
| 168 | 17Networks_LH_DefaultB_Temp_2 | -54 | -2 | -30 |
| 169 | 17Networks_LH_DefaultB_Temp_3 | -62 | -18 | -21 |
| 170 | 17Networks_LH_DefaultB_Temp_4 | -57 | -9 | -14 |
| 171 | 17Networks_LH_DefaultB_Temp_5 | -61 | -35 | -3 |
| 172 | 17Networks_LH_DefaultB_Temp_6 | -52 | -22 | -6 |
| 173 | 17Networks_LH_DefaultB_IPL_1 | -45 | -58 | 21 |
| 174 | 17Networks_LH_DefaultB_IPL_2 | -57 | -55 | 30 |
| 175 | 17Networks_LH_DefaultB_PFCd_1 | -4 | 51 | 28 |
| 176 | 17Networks_LH_DefaultB_PFCd_2 | -14 | 58 | 31 |
| 177 | 17Networks_LH_DefaultB_PFCd_3 | -22 | 51 | 31 |
| 178 | 17Networks_LH_DefaultB_PFCd_4 | -8 | 43 | 51 |
| 179 | 17Networks_LH_DefaultB_PFCd_5 | -13 | 24 | 61 |
| 180 | 17Networks_LH_DefaultB_PFCd_6 | -6 | 10 | 65 |
| 181 | 17Networks_LH_DefaultB_PFCl_1 | -41 | 19 | 48 |
| 182 | 17Networks_LH_DefaultB_PFCl_2 | -42 | 7 | 48 |
| 183 | 17Networks_LH_DefaultB_PFCv_1 | -36 | 22 | -16 |
| 184 | 17Networks_LH_DefaultB_PFCv_2 | -36 | 37 | -13 |
| 185 | 17Networks_LH_DefaultB_PFCv_3 | -46 | 32 | -10 |
| 186 | 17Networks_LH_DefaultB_PFCv_4 | -48 | 28 | 0 |
| 187 | 17Networks_LH_DefaultB_PFCv_5 | -53 | 19 | 11 |
| 188 | 17Networks_LH_DefaultC_IPL_1 | -40 | -79 | 30 |
| 189 | 17Networks_LH_DefaultC_Rsp_1 | -13 | -49 | 4 |
| 190 | 17Networks_LH_DefaultC_Rsp_2 | -8 | -52 | 9 |
| 191 | 17Networks_LH_DefaultC_Rsp_3 | -13 | -61 | 19 |
| 192 | 17Networks_LH_DefaultC_PHC_1 | -21 | -21 | -26 |
| 193 | 17Networks_LH_DefaultC_PHC_2 | -30 | -33 | -18 |
| 194 | 17Networks_LH_DefaultC_PHC_3 | -18 | -37 | -12 |
| 313 | 17Networks_RH_LimbicB_OFC_1 | 13 | 24 | -21 |
| 314 | 17Networks_RH_LimbicB_OFC_2 | 23 | 22 | -21 |
| 315 | 17Networks_RH_LimbicB_OFC_3 | 8 | 47 | -23 |
| 316 | 17Networks_RH_LimbicB_OFC_4 | 20 | 43 | -18 |
| 317 | 17Networks_RH_LimbicB_OFC_5 | 5 | 22 | -21 |
| 318 | 17Networks_RH_LimbicB_OFC_6 | 9 | 63 | -14 |
| 319 | 17Networks_RH_LimbicA_TempPole_1 | 28 | -1 | -40 |
| 320 | 17Networks_RH_LimbicA_TempPole_2 | 49 | -7 | -39 |
| 321 | 17Networks_RH_LimbicA_TempPole_3 | 37 | 17 | -38 |
| 322 | 17Networks_RH_LimbicA_TempPole_4 | 39 | -15 | -31 |
| 323 | 17Networks_RH_LimbicA_TempPole_5 | 29 | 12 | -30 |
| 324 | 17Networks_RH_LimbicA_TempPole_6 | 50 | -28 | -26 |
| 325 | 17Networks_RH_ContA_IPS_1 | 35 | -71 | 47 |
| 326 | 17Networks_RH_ContA_IPS_2 | 54 | -33 | 51 |
| 327 | 17Networks_RH_ContA_IPS_3 | 47 | -44 | 46 |
| 328 | 17Networks_RH_ContA_IPS_4 | 36 | -44 | 45 |
| 329 | 17Networks_RH_ContA_PFCd_1 | 24 | 10 | 58 |
| 330 | 17Networks_RH_ContA_PFCl_1 | 50 | 30 | 18 |
| 331 | 17Networks_RH_ContA_PFCl_2 | 48 | 18 | 23 |
| 332 | 17Networks_RH_ContA_PFCl_3 | 47 | 29 | 28 |
| 333 | 17Networks_RH_ContA_PFCl_4 | 49 | 8 | 25 |
| 334 | 17Networks_RH_ContA_PFCl_5 | 39 | 11 | 34 |
| 335 | 17Networks_RH_ContA_Cingm_1 | 5 | 1 | 30 |
| 336 | 17Networks_RH_ContB_Temp_1 | 62 | -28 | -20 |
| 337 | 17Networks_RH_ContB_Temp_2 | 63 | -42 | -11 |
| 338 | 17Networks_RH_ContB_IPL_1 | 55 | -45 | 33 |
| 339 | 17Networks_RH_ContB_IPL_2 | 54 | -53 | 44 |
| 340 | 17Networks_RH_ContB_IPL_3 | 56 | -41 | 48 |
| 341 | 17Networks_RH_ContB_IPL_4 | 41 | -55 | 48 |
| 342 | 17Networks_RH_ContB_PFCld_1 | 39 | 33 | 38 |
| 343 | 17Networks_RH_ContB_PFCld_2 | 45 | 19 | 44 |
| 344 | 17Networks_RH_ContB_PFCld_3 | 43 | 7 | 51 |
| 345 | 17Networks_RH_ContB_PFCld_4 | 34 | 15 | 56 |
| 346 | 17Networks_RH_ContB_PFClv_1 | 35 | 38 | -13 |
| 347 | 17Networks_RH_ContB_PFClv_2 | 28 | 55 | -14 |
| 348 | 17Networks_RH_ContB_PFClv_3 | 42 | 51 | -6 |
| 349 | 17Networks_RH_ContB_PFClv_4 | 27 | 59 | 3 |
| 350 | 17Networks_RH_ContB_PFCmp_1 | 5 | 28 | 48 |
| 351 | 17Networks_RH_ContC_pCun_1 | 17 | -63 | 28 |
| 352 | 17Networks_RH_ContC_pCun_2 | 13 | -71 | 39 |
| 353 | 17Networks_RH_ContC_pCun_3 | 5 | -64 | 44 |
| 354 | 17Networks_RH_ContC_pCun_4 | 7 | -50 | 45 |
| 355 | 17Networks_RH_ContC_pCun_5 | 8 | -71 | 53 |
| 356 | 17Networks_RH_ContC_Cingp_1 | 7 | -44 | 20 |
| 357 | 17Networks_RH_ContC_Cingp_2 | 6 | -26 | 28 |
| 358 | 17Networks_RH_DefaultA_Temp_1 | 61 | -8 | -23 |
| 359 | 17Networks_RH_DefaultA_IPL_1 | 53 | -53 | 26 |
| 360 | 17Networks_RH_DefaultA_IPL_2 | 47 | -64 | 42 |
| 361 | 17Networks_RH_DefaultA_PFCd_1 | 26 | 34 | 39 |
| 362 | 17Networks_RH_DefaultA_PFCd_2 | 24 | 26 | 51 |
| 363 | 17Networks_RH_DefaultA_pCunPCC_1 | 6 | -52 | 23 |
| 364 | 17Networks_RH_DefaultA_pCunPCC_2 | 5 | -63 | 31 |
| 365 | 17Networks_RH_DefaultA_pCunPCC_3 | 7 | -39 | 35 |
| 366 | 17Networks_RH_DefaultA_pCunPCC_4 | 4 | -20 | 37 |
| 367 | 17Networks_RH_DefaultA_pCunPCC_5 | 10 | -53 | 35 |
| 368 | 17Networks_RH_DefaultA_PFCm_1 | 5 | 41 | -11 |
| 369 | 17Networks_RH_DefaultA_PFCm_2 | 9 | 67 | 1 |
| 370 | 17Networks_RH_DefaultA_PFCm_3 | 7 | 42 | 4 |
| 371 | 17Networks_RH_DefaultA_PFCm_4 | 7 | 54 | 13 |
| 372 | 17Networks_RH_DefaultA_PFCm_5 | 17 | 65 | 16 |
| 373 | 17Networks_RH_DefaultA_PFCm_6 | 6 | 25 | 18 |
| 374 | 17Networks_RH_DefaultB_Temp_1 | 63 | -23 | -7 |
| 375 | 17Networks_RH_DefaultB_Temp_2 | 63 | -38 | 0 |
| 376 | 17Networks_RH_DefaultB_AntTemp_1 | 49 | 9 | -33 |
| 377 | 17Networks_RH_DefaultB_PFCd_1 | 6 | 58 | 29 |
| 378 | 17Networks_RH_DefaultB_PFCd_2 | 16 | 52 | 36 |
| 379 | 17Networks_RH_DefaultB_PFCd_3 | 5 | 44 | 40 |
| 380 | 17Networks_RH_DefaultB_PFCd_4 | 14 | 39 | 52 |
| 381 | 17Networks_RH_DefaultB_PFCd_5 | 12 | 20 | 63 |
| 382 | 17Networks_RH_DefaultB_PFCv_1 | 35 | 23 | -18 |
| 383 | 17Networks_RH_DefaultB_PFCv_2 | 48 | 32 | -8 |
| 384 | 17Networks_RH_DefaultB_PFCv_3 | 54 | 24 | 6 |
| 385 | 17Networks_RH_DefaultC_IPL_1 | 48 | -64 | 22 |
| 386 | 17Networks_RH_DefaultC_IPL_2 | 45 | -75 | 31 |
| 387 | 17Networks_RH_DefaultC_Rsp_1 | 14 | -46 | 4 |
| 388 | 17Networks_RH_DefaultC_Rsp_2 | 12 | -55 | 15 |
| 389 | 17Networks_RH_DefaultC_PHC_1 | 23 | -18 | -27 |
| 390 | 17Networks_RH_DefaultC_PHC_2 | 31 | -31 | -18 |
表S3 八个网络的所有节点信息
| 脑模板中的节点编号 | 节点名称 | 坐标 | ||
|---|---|---|---|---|
| R | A | S | ||
| 109 | 17Networks_LH_LimbicB_OFC_1 | -12 | 25 | -21 |
| 110 | 17Networks_LH_LimbicB_OFC_2 | -24 | 23 | -20 |
| 111 | 17Networks_LH_LimbicB_OFC_3 | -10 | 47 | -21 |
| 112 | 17Networks_LH_LimbicB_OFC_4 | -4 | 23 | -19 |
| 113 | 17Networks_LH_LimbicB_OFC_5 | -15 | 65 | -8 |
| 114 | 17Networks_LH_LimbicA_TempPole_1 | -37 | -5 | -42 |
| 115 | 17Networks_LH_LimbicA_TempPole_2 | -25 | 6 | -39 |
| 116 | 17Networks_LH_LimbicA_TempPole_3 | -26 | -9 | -33 |
| 117 | 17Networks_LH_LimbicA_TempPole_4 | -54 | -21 | -31 |
| 118 | 17Networks_LH_LimbicA_TempPole_5 | -40 | -21 | -27 |
| 119 | 17Networks_LH_LimbicA_TempPole_6 | -32 | 12 | -29 |
| 120 | 17Networks_LH_LimbicA_TempPole_7 | -44 | 5 | -17 |
| 121 | 17Networks_LH_ContA_Temp_1 | -55 | -62 | -1 |
| 122 | 17Networks_LH_ContA_IPS_1 | -29 | -74 | 42 |
| 123 | 17Networks_LH_ContA_IPS_2 | -58 | -42 | 45 |
| 124 | 17Networks_LH_ContA_IPS_3 | -35 | -62 | 48 |
| 125 | 17Networks_LH_ContA_IPS_4 | -45 | -41 | 47 |
| 126 | 17Networks_LH_ContA_IPS_5 | -33 | -46 | 41 |
| 127 | 17Networks_LH_ContA_PFCd_1 | -21 | 5 | 65 |
| 128 | 17Networks_LH_ContA_PFClv_1 | -48 | 35 | 10 |
| 129 | 17Networks_LH_ContA_PFClv_2 | -42 | 38 | 22 |
| 130 | 17Networks_LH_ContA_PFCl_1 | -49 | 6 | 26 |
| 131 | 17Networks_LH_ContA_PFCl_2 | -45 | 20 | 27 |
| 132 | 17Networks_LH_ContA_PFCl_3 | -39 | 7 | 34 |
| 133 | 17Networks_LH_ContA_Cingm_1 | -3 | 5 | 29 |
| 134 | 17Networks_LH_ContB_Temp_1 | -60 | -36 | -18 |
| 135 | 17Networks_LH_ContB_Temp_2 | -60 | -49 | -10 |
| 136 | 17Networks_LH_ContB_IPL_1 | -49 | -60 | 47 |
| 137 | 17Networks_LH_ContB_IPL_2 | -53 | -50 | 45 |
| 138 | 17Networks_LH_ContB_IPL_3 | -42 | -52 | 49 |
| 139 | 17Networks_LH_ContB_PFCd_1 | -30 | 14 | 57 |
| 140 | 17Networks_LH_ContB_PFClv_1 | -42 | 49 | -6 |
| 141 | 17Networks_LH_ContB_PFClv_2 | -28 | 58 | -1 |
| 142 | 17Networks_LH_ContB_PFClv_3 | -28 | 57 | 13 |
| 143 | 17Networks_LH_ContB_PFCmp_1 | -4 | 28 | 47 |
| 144 | 17Networks_LH_ContC_pCun_1 | -10 | -70 | 32 |
| 145 | 17Networks_LH_ContC_pCun_2 | -9 | -77 | 45 |
| 146 | 17Networks_LH_ContC_pCun_3 | -5 | -64 | 52 |
| 147 | 17Networks_LH_ContC_Cingp_1 | -6 | -41 | 24 |
| 148 | 17Networks_LH_ContC_Cingp_2 | -4 | -22 | 29 |
| 149 | 17Networks_LH_DefaultA_IPL_1 | -47 | -64 | 31 |
| 150 | 17Networks_LH_DefaultA_IPL_2 | -41 | -72 | 43 |
| 151 | 17Networks_LH_DefaultA_PFCd_1 | -25 | 28 | 43 |
| 152 | 17Networks_LH_DefaultA_PFCd_2 | -18 | 36 | 48 |
| 153 | 17Networks_LH_DefaultA_PFCd_3 | -22 | 20 | 52 |
| 154 | 17Networks_LH_DefaultA_pCunPCC_1 | -4 | -53 | 20 |
| 155 | 17Networks_LH_DefaultA_pCunPCC_2 | -5 | -60 | 30 |
| 156 | 17Networks_LH_DefaultA_pCunPCC_3 | -7 | -44 | 32 |
| 157 | 17Networks_LH_DefaultA_pCunPCC_4 | -4 | -34 | 38 |
| 158 | 17Networks_LH_DefaultA_pCunPCC_5 | -3 | -15 | 37 |
| 159 | 17Networks_LH_DefaultA_pCunPCC_6 | -3 | -68 | 41 |
| 160 | 17Networks_LH_DefaultA_pCunPCC_7 | -7 | -51 | 43 |
| 161 | 17Networks_LH_DefaultA_PFCm_1 | -5 | 55 | -10 |
| 162 | 17Networks_LH_DefaultA_PFCm_2 | -6 | 35 | -9 |
| 163 | 17Networks_LH_DefaultA_PFCm_3 | -6 | 59 | 7 |
| 164 | 17Networks_LH_DefaultA_PFCm_4 | -6 | 45 | 6 |
| 165 | 17Networks_LH_DefaultA_PFCm_5 | -16 | 67 | 8 |
| 166 | 17Networks_LH_DefaultA_PFCm_6 | -5 | 34 | 21 |
| 167 | 17Networks_LH_DefaultB_Temp_1 | -44 | 13 | -34 |
| 168 | 17Networks_LH_DefaultB_Temp_2 | -54 | -2 | -30 |
| 169 | 17Networks_LH_DefaultB_Temp_3 | -62 | -18 | -21 |
| 170 | 17Networks_LH_DefaultB_Temp_4 | -57 | -9 | -14 |
| 171 | 17Networks_LH_DefaultB_Temp_5 | -61 | -35 | -3 |
| 172 | 17Networks_LH_DefaultB_Temp_6 | -52 | -22 | -6 |
| 173 | 17Networks_LH_DefaultB_IPL_1 | -45 | -58 | 21 |
| 174 | 17Networks_LH_DefaultB_IPL_2 | -57 | -55 | 30 |
| 175 | 17Networks_LH_DefaultB_PFCd_1 | -4 | 51 | 28 |
| 176 | 17Networks_LH_DefaultB_PFCd_2 | -14 | 58 | 31 |
| 177 | 17Networks_LH_DefaultB_PFCd_3 | -22 | 51 | 31 |
| 178 | 17Networks_LH_DefaultB_PFCd_4 | -8 | 43 | 51 |
| 179 | 17Networks_LH_DefaultB_PFCd_5 | -13 | 24 | 61 |
| 180 | 17Networks_LH_DefaultB_PFCd_6 | -6 | 10 | 65 |
| 181 | 17Networks_LH_DefaultB_PFCl_1 | -41 | 19 | 48 |
| 182 | 17Networks_LH_DefaultB_PFCl_2 | -42 | 7 | 48 |
| 183 | 17Networks_LH_DefaultB_PFCv_1 | -36 | 22 | -16 |
| 184 | 17Networks_LH_DefaultB_PFCv_2 | -36 | 37 | -13 |
| 185 | 17Networks_LH_DefaultB_PFCv_3 | -46 | 32 | -10 |
| 186 | 17Networks_LH_DefaultB_PFCv_4 | -48 | 28 | 0 |
| 187 | 17Networks_LH_DefaultB_PFCv_5 | -53 | 19 | 11 |
| 188 | 17Networks_LH_DefaultC_IPL_1 | -40 | -79 | 30 |
| 189 | 17Networks_LH_DefaultC_Rsp_1 | -13 | -49 | 4 |
| 190 | 17Networks_LH_DefaultC_Rsp_2 | -8 | -52 | 9 |
| 191 | 17Networks_LH_DefaultC_Rsp_3 | -13 | -61 | 19 |
| 192 | 17Networks_LH_DefaultC_PHC_1 | -21 | -21 | -26 |
| 193 | 17Networks_LH_DefaultC_PHC_2 | -30 | -33 | -18 |
| 194 | 17Networks_LH_DefaultC_PHC_3 | -18 | -37 | -12 |
| 313 | 17Networks_RH_LimbicB_OFC_1 | 13 | 24 | -21 |
| 314 | 17Networks_RH_LimbicB_OFC_2 | 23 | 22 | -21 |
| 315 | 17Networks_RH_LimbicB_OFC_3 | 8 | 47 | -23 |
| 316 | 17Networks_RH_LimbicB_OFC_4 | 20 | 43 | -18 |
| 317 | 17Networks_RH_LimbicB_OFC_5 | 5 | 22 | -21 |
| 318 | 17Networks_RH_LimbicB_OFC_6 | 9 | 63 | -14 |
| 319 | 17Networks_RH_LimbicA_TempPole_1 | 28 | -1 | -40 |
| 320 | 17Networks_RH_LimbicA_TempPole_2 | 49 | -7 | -39 |
| 321 | 17Networks_RH_LimbicA_TempPole_3 | 37 | 17 | -38 |
| 322 | 17Networks_RH_LimbicA_TempPole_4 | 39 | -15 | -31 |
| 323 | 17Networks_RH_LimbicA_TempPole_5 | 29 | 12 | -30 |
| 324 | 17Networks_RH_LimbicA_TempPole_6 | 50 | -28 | -26 |
| 325 | 17Networks_RH_ContA_IPS_1 | 35 | -71 | 47 |
| 326 | 17Networks_RH_ContA_IPS_2 | 54 | -33 | 51 |
| 327 | 17Networks_RH_ContA_IPS_3 | 47 | -44 | 46 |
| 328 | 17Networks_RH_ContA_IPS_4 | 36 | -44 | 45 |
| 329 | 17Networks_RH_ContA_PFCd_1 | 24 | 10 | 58 |
| 330 | 17Networks_RH_ContA_PFCl_1 | 50 | 30 | 18 |
| 331 | 17Networks_RH_ContA_PFCl_2 | 48 | 18 | 23 |
| 332 | 17Networks_RH_ContA_PFCl_3 | 47 | 29 | 28 |
| 333 | 17Networks_RH_ContA_PFCl_4 | 49 | 8 | 25 |
| 334 | 17Networks_RH_ContA_PFCl_5 | 39 | 11 | 34 |
| 335 | 17Networks_RH_ContA_Cingm_1 | 5 | 1 | 30 |
| 336 | 17Networks_RH_ContB_Temp_1 | 62 | -28 | -20 |
| 337 | 17Networks_RH_ContB_Temp_2 | 63 | -42 | -11 |
| 338 | 17Networks_RH_ContB_IPL_1 | 55 | -45 | 33 |
| 339 | 17Networks_RH_ContB_IPL_2 | 54 | -53 | 44 |
| 340 | 17Networks_RH_ContB_IPL_3 | 56 | -41 | 48 |
| 341 | 17Networks_RH_ContB_IPL_4 | 41 | -55 | 48 |
| 342 | 17Networks_RH_ContB_PFCld_1 | 39 | 33 | 38 |
| 343 | 17Networks_RH_ContB_PFCld_2 | 45 | 19 | 44 |
| 344 | 17Networks_RH_ContB_PFCld_3 | 43 | 7 | 51 |
| 345 | 17Networks_RH_ContB_PFCld_4 | 34 | 15 | 56 |
| 346 | 17Networks_RH_ContB_PFClv_1 | 35 | 38 | -13 |
| 347 | 17Networks_RH_ContB_PFClv_2 | 28 | 55 | -14 |
| 348 | 17Networks_RH_ContB_PFClv_3 | 42 | 51 | -6 |
| 349 | 17Networks_RH_ContB_PFClv_4 | 27 | 59 | 3 |
| 350 | 17Networks_RH_ContB_PFCmp_1 | 5 | 28 | 48 |
| 351 | 17Networks_RH_ContC_pCun_1 | 17 | -63 | 28 |
| 352 | 17Networks_RH_ContC_pCun_2 | 13 | -71 | 39 |
| 353 | 17Networks_RH_ContC_pCun_3 | 5 | -64 | 44 |
| 354 | 17Networks_RH_ContC_pCun_4 | 7 | -50 | 45 |
| 355 | 17Networks_RH_ContC_pCun_5 | 8 | -71 | 53 |
| 356 | 17Networks_RH_ContC_Cingp_1 | 7 | -44 | 20 |
| 357 | 17Networks_RH_ContC_Cingp_2 | 6 | -26 | 28 |
| 358 | 17Networks_RH_DefaultA_Temp_1 | 61 | -8 | -23 |
| 359 | 17Networks_RH_DefaultA_IPL_1 | 53 | -53 | 26 |
| 360 | 17Networks_RH_DefaultA_IPL_2 | 47 | -64 | 42 |
| 361 | 17Networks_RH_DefaultA_PFCd_1 | 26 | 34 | 39 |
| 362 | 17Networks_RH_DefaultA_PFCd_2 | 24 | 26 | 51 |
| 363 | 17Networks_RH_DefaultA_pCunPCC_1 | 6 | -52 | 23 |
| 364 | 17Networks_RH_DefaultA_pCunPCC_2 | 5 | -63 | 31 |
| 365 | 17Networks_RH_DefaultA_pCunPCC_3 | 7 | -39 | 35 |
| 366 | 17Networks_RH_DefaultA_pCunPCC_4 | 4 | -20 | 37 |
| 367 | 17Networks_RH_DefaultA_pCunPCC_5 | 10 | -53 | 35 |
| 368 | 17Networks_RH_DefaultA_PFCm_1 | 5 | 41 | -11 |
| 369 | 17Networks_RH_DefaultA_PFCm_2 | 9 | 67 | 1 |
| 370 | 17Networks_RH_DefaultA_PFCm_3 | 7 | 42 | 4 |
| 371 | 17Networks_RH_DefaultA_PFCm_4 | 7 | 54 | 13 |
| 372 | 17Networks_RH_DefaultA_PFCm_5 | 17 | 65 | 16 |
| 373 | 17Networks_RH_DefaultA_PFCm_6 | 6 | 25 | 18 |
| 374 | 17Networks_RH_DefaultB_Temp_1 | 63 | -23 | -7 |
| 375 | 17Networks_RH_DefaultB_Temp_2 | 63 | -38 | 0 |
| 376 | 17Networks_RH_DefaultB_AntTemp_1 | 49 | 9 | -33 |
| 377 | 17Networks_RH_DefaultB_PFCd_1 | 6 | 58 | 29 |
| 378 | 17Networks_RH_DefaultB_PFCd_2 | 16 | 52 | 36 |
| 379 | 17Networks_RH_DefaultB_PFCd_3 | 5 | 44 | 40 |
| 380 | 17Networks_RH_DefaultB_PFCd_4 | 14 | 39 | 52 |
| 381 | 17Networks_RH_DefaultB_PFCd_5 | 12 | 20 | 63 |
| 382 | 17Networks_RH_DefaultB_PFCv_1 | 35 | 23 | -18 |
| 383 | 17Networks_RH_DefaultB_PFCv_2 | 48 | 32 | -8 |
| 384 | 17Networks_RH_DefaultB_PFCv_3 | 54 | 24 | 6 |
| 385 | 17Networks_RH_DefaultC_IPL_1 | 48 | -64 | 22 |
| 386 | 17Networks_RH_DefaultC_IPL_2 | 45 | -75 | 31 |
| 387 | 17Networks_RH_DefaultC_Rsp_1 | 14 | -46 | 4 |
| 388 | 17Networks_RH_DefaultC_Rsp_2 | 12 | -55 | 15 |
| 389 | 17Networks_RH_DefaultC_PHC_1 | 23 | -18 | -27 |
| 390 | 17Networks_RH_DefaultC_PHC_2 | 31 | -31 | -18 |
| 缩写 | 全称 | 脑区 |
|---|---|---|
| AntTemp | anterior temporal | 颞前部 |
| Cingm | mid-cingulate | 中扣带 |
| Cingp | cingulate posterior | 扣带后回 |
| IPL | inferior parietal lobule | 顶下小叶 |
| IPS | intraparietal sulcus | 顶内沟 |
| OFC | orbital frontal cortex | 眶额皮质 |
| pCun | precuneus | 楔前叶 |
| pCunPCC | precuneus posterior cingulate cortex | 楔前叶后扣带皮层 |
| PFCd | dorsal prefrontal cortex | 背侧前额叶 |
| PFCl | lateral prefrontal cortex | 外侧前额叶 |
| PFClv | lateral ventral prefrontal cortex | 腹外侧前额叶 |
| PFCm | medial prefrontal cortex | 中部前额叶 |
| PFCmp | medial posterior prefrontal cortex | 中后部前额叶 |
| PFCv | ventral prefrontal cortex | 腹侧前额叶 |
| PHC | parahippocampal cortex | 海马旁回 |
| Rsp | retrosplenial | 压后皮层 |
| Temp | temporal | 颞叶 |
| TempPole | temporal pole | 颞极 |
脑区缩写的标注:
| 缩写 | 全称 | 脑区 |
|---|---|---|
| AntTemp | anterior temporal | 颞前部 |
| Cingm | mid-cingulate | 中扣带 |
| Cingp | cingulate posterior | 扣带后回 |
| IPL | inferior parietal lobule | 顶下小叶 |
| IPS | intraparietal sulcus | 顶内沟 |
| OFC | orbital frontal cortex | 眶额皮质 |
| pCun | precuneus | 楔前叶 |
| pCunPCC | precuneus posterior cingulate cortex | 楔前叶后扣带皮层 |
| PFCd | dorsal prefrontal cortex | 背侧前额叶 |
| PFCl | lateral prefrontal cortex | 外侧前额叶 |
| PFClv | lateral ventral prefrontal cortex | 腹外侧前额叶 |
| PFCm | medial prefrontal cortex | 中部前额叶 |
| PFCmp | medial posterior prefrontal cortex | 中后部前额叶 |
| PFCv | ventral prefrontal cortex | 腹侧前额叶 |
| PHC | parahippocampal cortex | 海马旁回 |
| Rsp | retrosplenial | 压后皮层 |
| Temp | temporal | 颞叶 |
| TempPole | temporal pole | 颞极 |
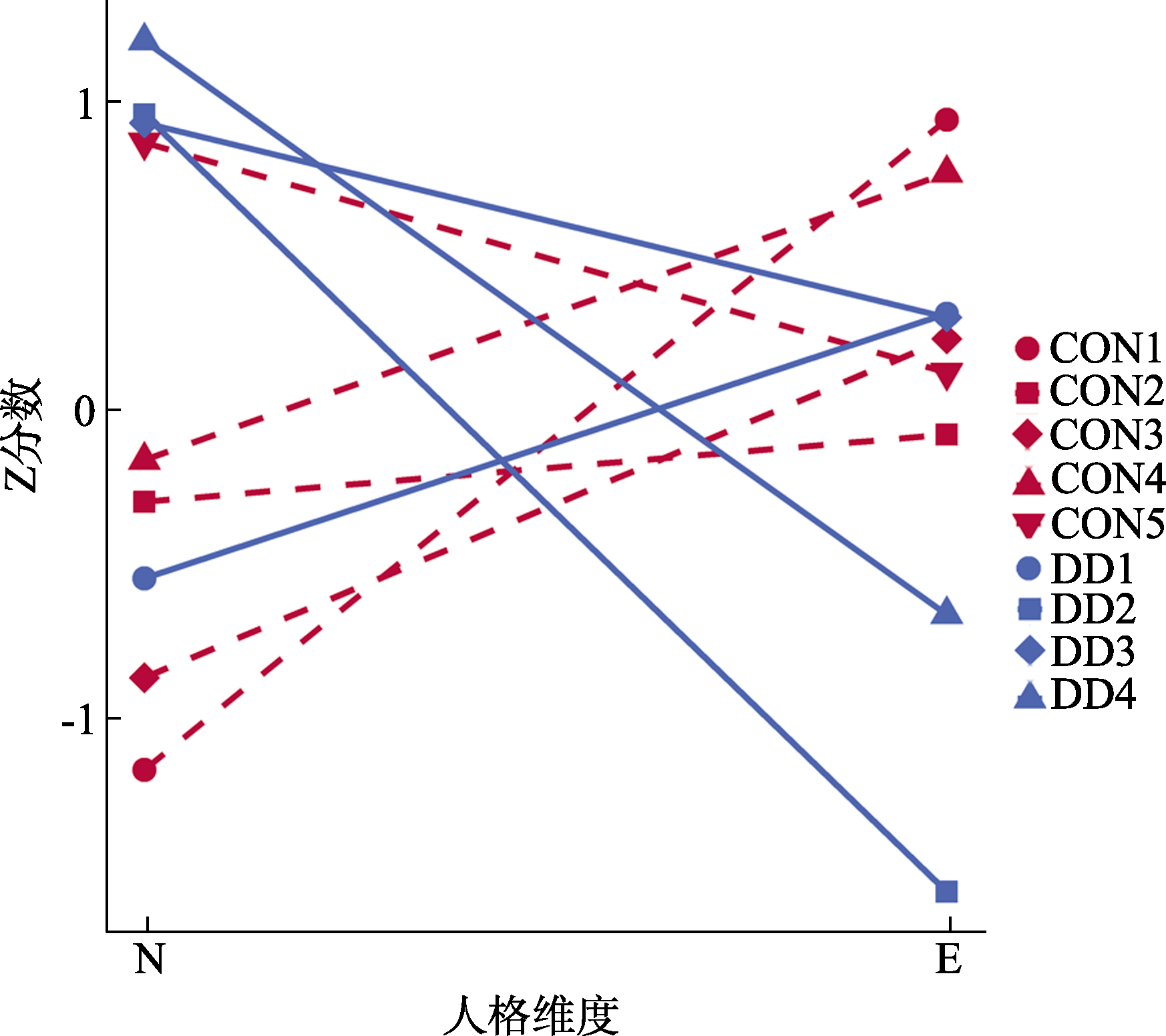
图2 抑郁症与控制组人格类型的神经质和外向性的Z分数 注:抑郁症类型1呈现低神经质和中等偏高的外向性水平; 类型2和4呈现出高神经质和低外向性水平, 类型3呈现出高神经质和中等偏高的外向性水平。控制组类型1与类型3呈现低神经质, 高或者中等偏高的外向性水平; 类型2的神经质和外向性都处于中等偏低的程度; 类型4呈现中等神经质和高外向性的趋势; 类型5则呈现高神经质和中等外向性的趋势。N: 神经质(neuroticism); E: 外向性(extraversion) ; CON: control控制组; DD: depressive disorder 抑郁症。DD1: 抑郁症类型1; CON1: 控制组类型1。
| 组间比较 | 左侧杏仁核−边缘网络 | 左侧脑岛−边缘网络 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| t | pholm | Cohen's d | 95% CI | t | pholm | Cohen's d | 95% CI | |||||
| DD1 | − | DD4 | −0.87 | 1.000 | −0.229 | −0.747 | 0.288 | −0.03 | 1.000 | −0.007 | −0.525 | 0.510 |
| − | DD3 | −1.82 | 0.841 | −0.467 | −0.974 | 0.041 | −1.78 | 1.000 | −0.456 | −0.963 | 0.052 | |
| − | DD2 | −0.48 | 1.000 | −0.126 | −0.639 | 0.387 | 1.16 | 1.000 | 0.301 | −0.212 | 0.815 | |
| − | CON2 | 0.75 | 1.000 | 0.204 | −0.336 | 0.744 | 0.96 | 1.000 | 0.263 | −0.278 | 0.803 | |
| − | CON1 | −2.78 | 0.099 | −0.689 | −1.181 | −0.197 | −1.60 | 1.000 | −0.396 | −0.886 | 0.093 | |
| − | CON3 | −3.47 | 0.013 | −0.977 | −1.540 | −0.413 | −2.60 | 0.159 | −0.733 | −1.292 | −0.173 | |
| DD4 | − | DD3 | −0.91 | 1.000 | −0.237 | −0.751 | 0.277 | −1.72 | 1.000 | −0.448 | −0.964 | 0.067 |
| − | DD2 | 0.40 | 1.000 | 0.103 | −0.412 | 0.619 | 1.18 | 1.000 | 0.309 | −0.207 | 0.825 | |
| − | CON2 | 1.46 | 1.000 | 0.433 | −0.153 | 1.020 | 0.91 | 1.000 | 0.270 | −0.316 | 0.855 | |
| − | CON1 | −1.75 | 0.892 | −0.460 | −0.978 | 0.059 | −1.48 | 1.000 | −0.389 | −0.908 | 0.129 | |
| − | CON3 | −2.72 | 0.114 | −0.747 | −1.294 | −0.201 | −2.64 | 0.155 | −0.725 | −1.272 | −0.179 | |
| DD3 | − | DD2 | 1.37 | 1.000 | 0.341 | −0.151 | 0.833 | 3.04 | 0.053 | 0.757 | 0.261 | 1.253 |
| − | CON2 | 2.24 | 0.390 | 0.671 | 0.077 | 1.264 | 2.40 | 0.241 | 0.718 | 0.125 | 1.312 | |
| − | CON1 | −0.86 | 1.000 | −0.222 | −0.735 | 0.290 | 0.23 | 1.000 | 0.059 | −0.453 | 0.571 | |
| − | CON3 | −2.00 | 0.613 | −0.510 | −1.016 | −0.004 | −1.08 | 1.000 | −0.277 | −0.781 | 0.227 | |
| DD2 | − | CON2 | 1.08 | 1.000 | 0.330 | −0.271 | 0.931 | −0.13 | 1.000 | −0.039 | −0.639 | 0.561 |
| − | CON1 | −2.14 | 0.470 | −0.563 | −1.085 | −0.041 | −2.65 | 0.155 | −0.698 | −1.221 | −0.175 | |
| − | CON3 | −3.34 | 0.018 | −0.851 | −1.359 | −0.342 | −4.06 | 0.001 | −1.034 | −1.545 | −0.523 | |
| CON2 | − | CON1 | −3.38 | 0.016 | −0.893 | −1.421 | −0.365 | −2.49 | 0.201 | −0.659 | −1.184 | −0.134 |
| − | CON3 | −3.49 | 0.013 | −1.180 | −1.858 | −0.503 | −2.94 | 0.070 | −0.995 | −1.670 | −0.321 | |
| CON1 | − | CON3 | −0.99 | 1.000 | −0.288 | −0.859 | 0.284 | −1.16 | 1.000 | −0.336 | −0.907 | 0.235 |
表2 左侧杏仁核/脑岛−边缘网络功能连接强度的事后检验结果
| 组间比较 | 左侧杏仁核−边缘网络 | 左侧脑岛−边缘网络 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| t | pholm | Cohen's d | 95% CI | t | pholm | Cohen's d | 95% CI | |||||
| DD1 | − | DD4 | −0.87 | 1.000 | −0.229 | −0.747 | 0.288 | −0.03 | 1.000 | −0.007 | −0.525 | 0.510 |
| − | DD3 | −1.82 | 0.841 | −0.467 | −0.974 | 0.041 | −1.78 | 1.000 | −0.456 | −0.963 | 0.052 | |
| − | DD2 | −0.48 | 1.000 | −0.126 | −0.639 | 0.387 | 1.16 | 1.000 | 0.301 | −0.212 | 0.815 | |
| − | CON2 | 0.75 | 1.000 | 0.204 | −0.336 | 0.744 | 0.96 | 1.000 | 0.263 | −0.278 | 0.803 | |
| − | CON1 | −2.78 | 0.099 | −0.689 | −1.181 | −0.197 | −1.60 | 1.000 | −0.396 | −0.886 | 0.093 | |
| − | CON3 | −3.47 | 0.013 | −0.977 | −1.540 | −0.413 | −2.60 | 0.159 | −0.733 | −1.292 | −0.173 | |
| DD4 | − | DD3 | −0.91 | 1.000 | −0.237 | −0.751 | 0.277 | −1.72 | 1.000 | −0.448 | −0.964 | 0.067 |
| − | DD2 | 0.40 | 1.000 | 0.103 | −0.412 | 0.619 | 1.18 | 1.000 | 0.309 | −0.207 | 0.825 | |
| − | CON2 | 1.46 | 1.000 | 0.433 | −0.153 | 1.020 | 0.91 | 1.000 | 0.270 | −0.316 | 0.855 | |
| − | CON1 | −1.75 | 0.892 | −0.460 | −0.978 | 0.059 | −1.48 | 1.000 | −0.389 | −0.908 | 0.129 | |
| − | CON3 | −2.72 | 0.114 | −0.747 | −1.294 | −0.201 | −2.64 | 0.155 | −0.725 | −1.272 | −0.179 | |
| DD3 | − | DD2 | 1.37 | 1.000 | 0.341 | −0.151 | 0.833 | 3.04 | 0.053 | 0.757 | 0.261 | 1.253 |
| − | CON2 | 2.24 | 0.390 | 0.671 | 0.077 | 1.264 | 2.40 | 0.241 | 0.718 | 0.125 | 1.312 | |
| − | CON1 | −0.86 | 1.000 | −0.222 | −0.735 | 0.290 | 0.23 | 1.000 | 0.059 | −0.453 | 0.571 | |
| − | CON3 | −2.00 | 0.613 | −0.510 | −1.016 | −0.004 | −1.08 | 1.000 | −0.277 | −0.781 | 0.227 | |
| DD2 | − | CON2 | 1.08 | 1.000 | 0.330 | −0.271 | 0.931 | −0.13 | 1.000 | −0.039 | −0.639 | 0.561 |
| − | CON1 | −2.14 | 0.470 | −0.563 | −1.085 | −0.041 | −2.65 | 0.155 | −0.698 | −1.221 | −0.175 | |
| − | CON3 | −3.34 | 0.018 | −0.851 | −1.359 | −0.342 | −4.06 | 0.001 | −1.034 | −1.545 | −0.523 | |
| CON2 | − | CON1 | −3.38 | 0.016 | −0.893 | −1.421 | −0.365 | −2.49 | 0.201 | −0.659 | −1.184 | −0.134 |
| − | CON3 | −3.49 | 0.013 | −1.180 | −1.858 | −0.503 | −2.94 | 0.070 | −0.995 | −1.670 | −0.321 | |
| CON1 | − | CON3 | −0.99 | 1.000 | −0.288 | −0.859 | 0.284 | −1.16 | 1.000 | −0.336 | −0.907 | 0.235 |
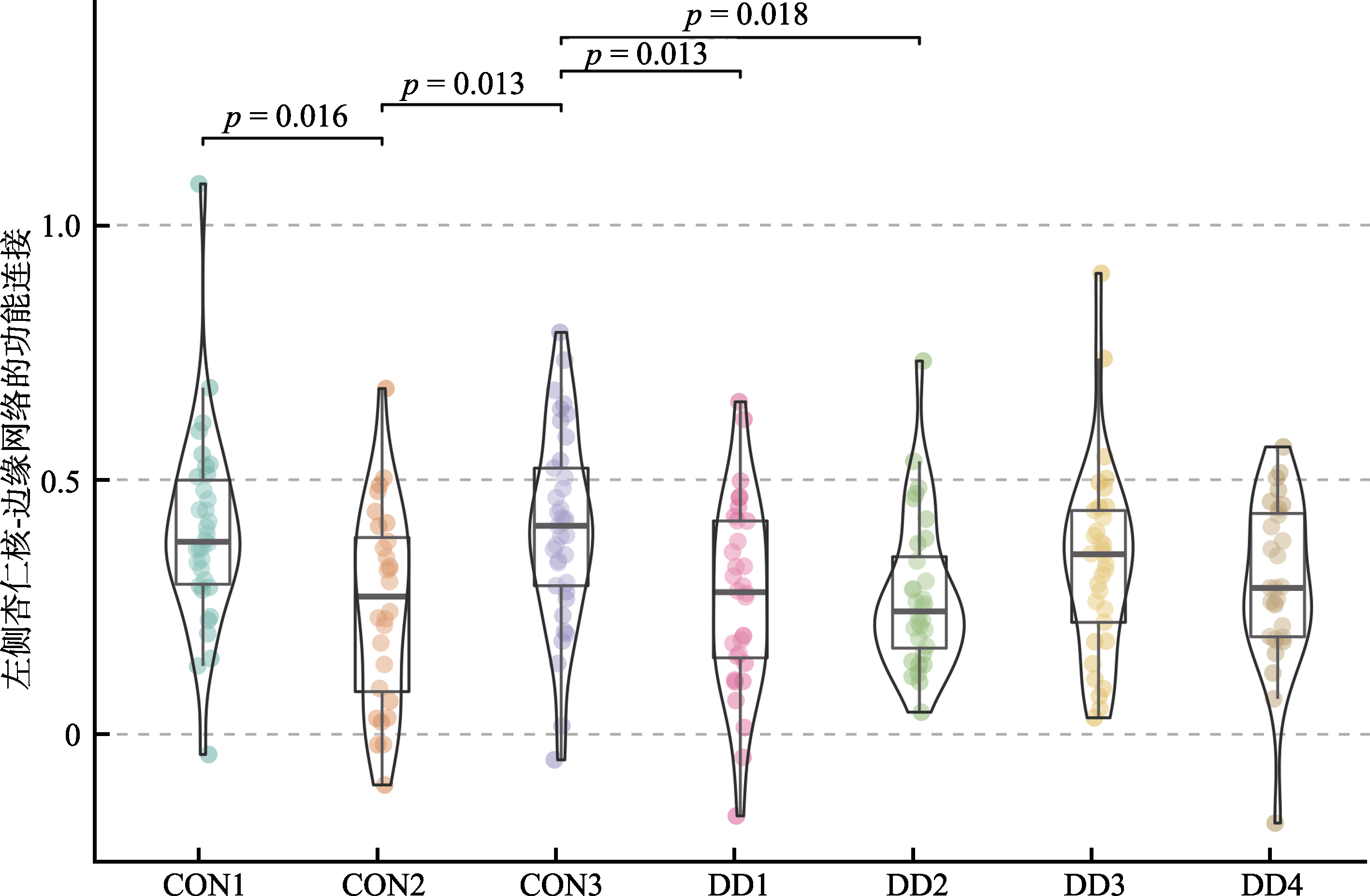
图3 左侧杏仁核与边缘网络的功能连接强度的组间差异 注:CON: control控制组; DD: depressive disorder 抑郁症。DD1: 抑郁症类型1; CON1: 控制组类型1。
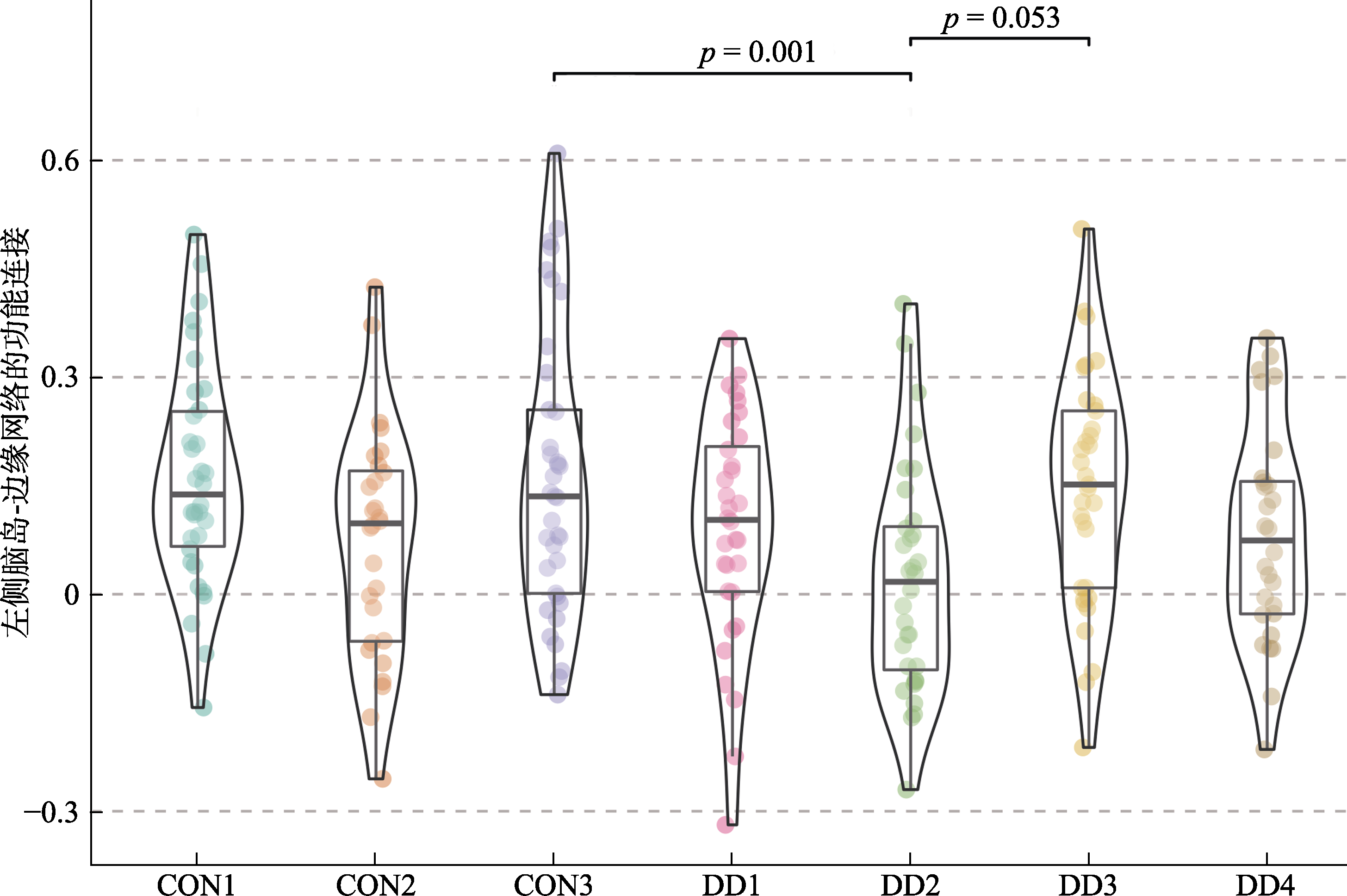
图4 左侧脑岛与边缘网络的功能连接强度的组间差异 注:CON: control控制组; DD: depressive disorder 抑郁症。DD1: 抑郁症类型1; CON1: 控制组类型1。
| [1] | Abbasi, M., Mirderikvand, F., Adavi, H., & Hojati, M. (2018). The relationship between personality traits (neuroticism and extraversion) and self-efficacy with aging depression. Iranian Journal of Ageing, 12(4), 458-466. |
| [2] |
Alexopoulos, G. S. (2002). Frontostriatal and limbic dysfunction in late-life depression. The American Journal of Geriatric Psychiatry, 10(6), 687-695.
doi: 10.1097/00019442-200211000-00007 URL |
| [3] |
Allen, T. A., Carey, B. E., McBride, C., Bagby, R. M., deYoung, C. G., & Quilty, L. C. (2018). Big Five aspects of personality interact to predict depression. Journal of Personality, 86(4), 714-725.
doi: 10.1111/jopy.12352 pmid: 28921930 |
| [4] |
Altman, N., & Krzywinski, M. (2017). Points of significance: Clustering. Nature Methods, 14(6), 545-547.
doi: 10.1038/nmeth.4299 URL |
| [5] |
Asendorpf, J. B., Borkenau, P., Ostendorf, F., & van Aken, M. A. (2001). Carving personality description at its joints: Confirmation of three replicable personality prototypes for both children and adults. European Journal of Personality, 15(3), 169-198.
doi: 10.1002/per.408 URL |
| [6] |
Banjongrewadee, M., Wongpakaran, N., Wongpakaran, T., Pipanmekaporn, T., Punjasawadwong, Y., & Mueankwan, S. (2020). The role of perceived stress and cognitive function on the relationship between neuroticism and depression among the elderly: A structural equation model approach. BMC Psychiatry, 20(1), 1-8.
doi: 10.1186/s12888-019-2374-2 |
| [7] | Blondel, V. D., Guillaume, J. -L., Lambiotte, R., & Lefebvre, E. (2008). Fast unfolding of communities in large networks. Journal of Statistical Mechanics: Theory and Experiment, 2008(10), P10008. |
| [8] |
Bohane, L., Maguire, N., & Richardson, T. (2017). Resilients, overcontrollers and undercontrollers: A systematic review of the utility of a personality typology method in understanding adult mental health problems. Clinical Psychology Review, 57, 75-92.
doi: S0272-7358(16)30294-X pmid: 28850932 |
| [9] |
Bowden, S., Saklofske, D., van de Vijver, F., Sudarshan, N., & Eysenck, S. (2016). Cross-cultural measurement invariance of the Eysenck Personality Questionnaire across 33 countries. Personality and Individual Differences, 103, 53-60.
doi: 10.1016/j.paid.2016.04.028 URL |
| [10] |
Breiman, L. (2001). Random forests. Machine Learning, 45(1), 5-32.
doi: 10.1023/A:1010933404324 URL |
| [11] |
Bryan, C. J., Tipton, E., & Yeager, D. S. (2021). Behavioural science is unlikely to change the world without a heterogeneity revolution. Nature Human Behaviour, 5(8), 980-989.
doi: 10.1038/s41562-021-01143-3 pmid: 34294901 |
| [12] |
Cale, E. M. (2006). A quantitative review of the relations between the “Big 3” higher order personality dimensions and antisocial behavior. Journal of Research in Personality, 40(3), 250-284.
doi: 10.1016/j.jrp.2005.01.001 URL |
| [13] |
Canli, T. (2004). Functional brain mapping of extraversion and neuroticism: :Learning from individual differences in emotion processing. Journal of Personality, 72(6), 1105-1132.
doi: 10.1111/j.1467-6494.2004.00292.x URL |
| [14] |
Chand, G. B., Dwyer, D. B., Erus, G., Sotiras, A., Varol, E., Srinivasan, D., ... Dazzan, P. (2020). Two distinct neuroanatomical subtypes of schizophrenia revealed using machine learning. Brain, 143(3), 1027-1038.
doi: 10.1093/brain/awaa025 pmid: 32103250 |
| [15] | Chen, Z. -G. (1983). Item analysis of Eysenck Personality Questionnaire tested in Beijing-district. Acta Psychologica Sinica, 15(2), 85-92. |
| [陈仲庚. (1983). 艾森克人格问卷的项目分析. 心理学报, 15(2), 85-92.] | |
| [16] |
Cheng, W., Rolls, E. T., Qiu, J., Xie, X., Lyu, W., Li, Y., ... Feng, J. F. (2018). Functional connectivity of the human amygdala in health and in depression. Social Cognitive and Affective Neuroscience, 13(6), 557-568.
doi: 10.1093/scan/nsy032 pmid: 29767786 |
| [17] |
Cheng, W., Rolls, E. T., Qiu, J., Xie, X., Wei, D., Huang, C. -C., ... Feng, J. F. (2018). Increased functional connectivity of the posterior cingulate cortex with the lateral orbitofrontal cortex in depression. Translational Psychiatry, 8(1), 1-10.
doi: 10.1038/s41398-017-0025-2 |
| [18] |
Cohen, M. X., Young, J., Baek, J. -M., Kessler, C., & Ranganath, C. (2005). Individual differences in extraversion and dopamine genetics predict neural reward responses. Cognitive Brain Research, 25(3), 851-861.
pmid: 16289773 |
| [19] |
Cole, M. W., Bassett, D. S., Power, J. D., Braver, T. S., & Petersen, S. E. (2014). Intrinsic and task-evoked network architectures of the human brain. Neuron, 83(1), 238-251.
doi: 10.1016/j.neuron.2014.05.014 pmid: 24991964 |
| [20] |
Dollinger, S. J., Urban, K. K., & James, T. A. (2004). Creativity and openness: Further validation of two creative product measures. Creativity Research Journal, 16(1), 35-47.
doi: 10.1207/s15326934crj1601_4 URL |
| [21] |
Donnellan, M. B., & Robins, R. W. (2010). Resilient, overcontrolled, and undercontrolled personality types: Issues and controversies. Social and Personality Psychology Compass, 4(11), 1070-1083.
doi: 10.1111/j.1751-9004.2010.00313.x URL |
| [22] |
Duggan, C., Sham, P., Lee, A., Minne, C., & Murray, R. (1995). Neuroticism: A vulnerability marker for depression evidence from a family study. Journal of Affective Disorders, 35(3), 139-143.
pmid: 8749842 |
| [23] |
Etkin, A., Egner, T., & Kalisch, R. (2011). Emotional processing in anterior cingulate and medial prefrontal cortex. Trends in Cognitive Sciences, 15(2), 85-93.
doi: 10.1016/j.tics.2010.11.004 pmid: 21167765 |
| [24] |
Feczko, E., Miranda-Dominguez, O., Marr, M., Graham, A. M., Nigg, J. T., & Fair, D. A. (2019). The heterogeneity problem: Approaches to identify psychiatric subtypes. Trends in Cognitive Sciences, 23(7), 584-601.
doi: S1364-6613(19)30092-0 pmid: 31153774 |
| [25] | Fournier, J., Jones, N., Chase, H., Cummings, L., Graur, S., & Phillips, M. (2017). Personality dysfunction in depression and individual differences in effortful emotion regulation. Biological Psychiatry, 81(10), S336-S337. |
| [26] |
Friston, K. J., Williams, S., Howard, R., Frackowiak, R. S., & Turner, R. (1996). Movement-related effects in fMRI time-series. Magnetic Resonance in Medicine, 35(3), 346-355.
doi: 10.1002/mrm.1910350312 pmid: 8699946 |
| [27] |
Gerlach, M., Farb, B., Revelle, W., & Amaral, L. A. N. (2018). A robust data-driven approach identifies four personality types across four large data sets. Nature Human Behaviour, 2(10), 735-742.
doi: 10.1038/s41562-018-0419-z pmid: 31406291 |
| [28] |
Haas, B. W., Omura, K., Constable, R. T., & Canli, T. (2007). Emotional conflict and neuroticism: Personality-dependent activation in the amygdala and subgenual anterior cingulate. Behavioral Neuroscience, 121(2), 249-256.
pmid: 17469914 |
| [29] |
Hamilton, M. (1960). A rating scale for depression. Journal of Neurology, Neurosurgery, and Psychiatry, 23(1), 56-62.
doi: 10.1136/jnnp.23.1.56 URL |
| [30] |
Herzberg, P. Y., & Roth, M. (2006). Beyond resilients, undercontrollers, and overcontrollers? An extension of personality prototype research. European Journal of Personality, 20(1), 5-28.
doi: 10.1002/per.557 URL |
| [31] |
Hooker, C. I., Verosky, S. C., Miyakawa, A., Knight, R. T., & D’Esposito, M. (2008). The influence of personality on neural mechanisms of observational fear and reward learning. Neuropsychologia, 46(11), 2709-2724.
doi: 10.1016/j.neuropsychologia.2008.05.005 pmid: 18573512 |
| [32] |
Isler, L., Fletcher, G. J., Liu, J. H., & Sibley, C. G. (2017). Validation of the four-profile configuration of personality types within the Five-Factor Model. Personality and Individual Differences, 106, 257-262.
doi: 10.1016/j.paid.2016.10.058 URL |
| [33] |
Jones, L., Scott, J., Cooper, C., Forty, L., Smith, K. G., Sham, P., ... Jones, I. (2010). Cognitive style, personality and vulnerability to postnatal depression. British Journal of Psychiatry, 196(3), 200-205.
doi: 10.1192/bjp.bp.109.064683 URL |
| [34] |
Kehoe, E. G., Toomey, J. M., Balsters, J. H., & Bokde, A. L. (2012). Personality modulates the effects of emotional arousal and valence on brain activation. Social Cognitive and Affective Neuroscience, 7(7), 858-870.
doi: 10.1093/scan/nsr059 pmid: 21948954 |
| [35] |
Kendler, K. S., Kuhn, J., & Prescott, C. A. (2004). The interrelationship of neuroticism, sex, and stressful life events in the prediction of episodes of major depression. American Journal of Psychiatry, 161(4), 631-636.
doi: 10.1176/appi.ajp.161.4.631 pmid: 15056508 |
| [36] |
Kercher, A. J., Rapee, R. M., & Schniering, C. A. (2009). Neuroticism, life events and negative thoughts in the development of depression in adolescent girls. Journal of Abnormal Child Psychology, 37(7), 903-915.
doi: 10.1007/s10802-009-9325-1 pmid: 19437113 |
| [37] |
Khoo, S., & Simms, L. J. (2018). Links between depression and openness and its facets. Personality and Mental Health, 12(3), 203-215.
doi: 10.1002/pmh.1417 pmid: 29611346 |
| [38] |
Kim, M. J., Loucks, R. A., Palmer, A. L., Brown, A. C., Solomon, K. M., Marchante, A. N., & Whalen, P. J. (2011). The structural and functional connectivity of the amygdala: From normal emotion to pathological anxiety. Behavioural Brain Research, 223(2), 403-410.
doi: 10.1016/j.bbr.2011.04.025 pmid: 21536077 |
| [39] |
Kim, S. Y., Stewart, R., Bae, K. Y., Kim, S. W., Shin, I. S., Hong, Y. J., ... Kim, J. M. (2016). Influences of the Big Five personality traits on the treatment response and longitudinal course of depression in patients with acute coronary syndrome: A randomised controlled trial. Journal of Affective Disorders, 203, 38-45.
doi: 10.1016/j.jad.2016.05.071 URL |
| [40] |
Klein, D. N., Kotov, R., & Bufferd, S. J. (2011). Personality and depression: Explanatory models and review of the evidence. Annual Review of Clinical Psychology, 7, 269-295.
doi: 10.1146/annurev-clinpsy-032210-104540 pmid: 21166535 |
| [41] |
Knyazev, G. G. (2006). EEG correlates of personality types. Netherlands Journal of Psychology, 62(2), 78-87.
doi: 10.1007/BF03061054 URL |
| [42] |
Kokkonen, M., & Pulkkinen, L. (2001). Extraversion and neuroticism as antecedents of emotion regulation and dysregulation in adulthood. European Journal of Personality, 15(6), 407-424.
doi: 10.1002/per.425 URL |
| [43] |
Kong, R., Li, J., Orban, C., Sabuncu, M. R., Liu, H., Schaefer, A., ... Eickhoff, S. B. (2019). Spatial topography of individual- specific cortical networks predicts human cognition, personality, and emotion. Cerebral Cortex, 29(6), 2533-2551.
doi: 10.1093/cercor/bhy123 |
| [44] |
Leikas, S., & Salmela-Aro, K. (2014). Personality types during transition to young adulthood: How are they related to life situation and well-being? Journal of Adolescence, 37(5), 753-762.
doi: 10.1016/j.adolescence.2014.01.003 pmid: 24529315 |
| [45] |
Lorenzetti, V., Allen, N. B., Fornito, A., & Yücel, M. (2009). Structural brain abnormalities in major depressive disorder: a selective review of recent MRI studies. Journal of Affective Disorders, 117(1-2), 1-17.
doi: 10.1016/j.jad.2008.11.021 pmid: 19237202 |
| [46] |
Mantini, D., Corbetta, M., Romani, G. L., Orban, G. A., & Vanduffel, W. (2013). Evolutionarily novel functional networks in the human brain? Journal of Neuroscience, 33(8), 3259-3275.
doi: 10.1523/JNEUROSCI.4392-12.2013 pmid: 23426655 |
| [47] | McDonnell, S., & Semkovska, M. (2020). Resilience as mediator between extraversion, neuroticism, and depressive symptoms in university students. Journal of Positive School Psychology, 4(1), 26-40. |
| [48] |
Menon, V., & Uddin, L. Q. (2010). Saliency, switching, attention and control: A network model of insula function. Brain Structure and Function, 214(5-6), 655-667.
doi: 10.1007/s00429-010-0262-0 pmid: 20512370 |
| [49] | Miller, C., Brady, M., & Dean, S. (2020). Is Conscientiousness a proxy for Depression? A study of Big Five Con-scientiousness predictive power above depression. Journal of Mental Health and Social Behaviour, 2(1), 112. |
| [50] |
Milne, A. M., MacQueen, G. M., & Hall, G. B. (2012). Abnormal hippocampal activation in patients with extensive history of major depression: An fMRI study. Journal of Psychiatry and Neuroscience, 37(1), 28-36.
doi: 10.1503/jpn.110004 URL |
| [51] |
Mobbs, D., Hagan, C. C., Azim, E., Menon, V., & Reiss, A. L. (2005). Personality predicts activity in reward and emotional regions associated with humor. Proceedings of the National Academy of Sciences, 102(45), 16502-16506.
doi: 10.1073/pnas.0408457102 URL |
| [52] |
Murray, L. E., & O'Neill, L. (2019). Neuroticism and extraversion mediate the relationship between having a sibling with developmental disabilities and anxiety and depression symptoms. Journal of Affective Disorders, 243, 232-240.
doi: S0165-0327(18)30316-1 pmid: 30248634 |
| [53] |
Ormel, J., Bastiaansen, A., Riese, H., Bos, E. H., Servaas, M., Ellenbogen, M., ... Aleman, A. (2013). The biological and psychological basis of neuroticism: Current status and future directions. Neuroscience & Biobehavioral Reviews, 37(1), 59-72.
doi: 10.1016/j.neubiorev.2012.09.004 URL |
| [54] | Pang, Y. J. (2020). Brain connectivity study of depressive disorder and neuroticism personality (Unpublished doctorial dissertation). University of Electronic Science and Technology of China. |
| [庞雅静. (2020). 抑郁症和神经质人格的脑连接研究 (博士学位论文). 电子科技大学, 成都.] | |
| [55] |
Paulus, M. P., Rogalsky, C., Simmons, A., Feinstein, J. S., & Stein, M. B. (2003). Increased activation in the right insula during risk-taking decision making is related to harm avoidance and neuroticism. NeuroImage, 19(4), 1439-1448.
pmid: 12948701 |
| [56] |
Peluso, M. A., Glahn, D. C., Matsuo, K., Monkul, E. S., Najt, P., Zamarripa, F., ... Soaress, J. C. (2009). Amygdala hyperactivation in untreated depressed individuals. Psychiatry Research: Neuroimaging, 173(2), 158-161.
doi: 10.1016/j.pscychresns.2009.03.006 URL |
| [57] |
Peters, A. T., Burkhouse, K., Feldhaus, C. C., Langenecker, S. A., & Jacobs, R. H. (2016). Aberrant resting-state functional connectivity in limbic and cognitive control networks relates to depressive rumination and mindfulness: A pilot study among adolescents with a history of depression. Journal of Affective Disorders, 200, 178-181.
doi: 10.1016/j.jad.2016.03.059 pmid: 27136416 |
| [58] |
Power, J. D., Mitra, A., Laumann, T. O., Snyder, A. Z., Schlaggar, B. L., & Petersen, S. E. (2014). Methods to detect, characterize, and remove motion artifact in resting state fMRI. NeuroImage, 84, 320-341.
doi: 10.1016/j.neuroimage.2013.08.048 pmid: 23994314 |
| [59] |
Quirk, G. J., Russo, G. K., Barron, J. L., & Lebron, K. (2000). The role of ventromedial prefrontal cortex in the recovery of extinguished fear. Journal of Neuroscience, 20(16), 6225-6231.
pmid: 10934272 |
| [60] | Rai, S., Griffiths, K. R., Breukelaar, I. A., Barreiros, A. R., Chen, W., Boyce, P., ... Korgaonkar, M. S. (2021). Default- mode and fronto-parietal network connectivity during rest distinguishes asymptomatic patients with bipolar disorder and major depressive disorder. Translational Psychiatry, 11(1), 547. |
| [61] |
Rocca, M. A., Pravata, E., Valsasina, P., Radaelli, M., Colombo, B., Vacchi, L., ... Filippi, M. (2015). Hippocampal- DMN disconnectivity in MS is related to WM lesions and depression. Human Brain Mapping, 36(12), 5051-5063.
doi: 10.1002/hbm.v36.12 URL |
| [62] |
Rolls, E. T. (2015). Limbic systems for emotion and for memory, but no single limbic system. Cortex, 62, 119-157.
doi: 10.1016/j.cortex.2013.12.005 pmid: 24439664 |
| [63] |
Scalabrini, A., Vai, B., Poletti, S., Damiani, S., Mucci, C., Colombo, C., ... Northoff, G. (2020). All roads lead to the default-mode network-global source of DMN abnormalities in major depressive disorder. Neuropsychopharmacology, 45(12), 2058-2069.
doi: 10.1038/s41386-020-0785-x |
| [64] |
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X. -N., Holmes, A. J., ... Yeo, B. T. (2018). Local-global parcellation of the human cerebral cortex from intrinsic functional connectivity MRI. Cerebral Cortex, 28(9), 3095-3114.
doi: 10.1093/cercor/bhx179 URL |
| [65] |
Spielberg, J. M., Forbes, E. E., Ladouceur, C. D., Worthman, C. M., Olino, T. M., Ryan, N. D., & Dahl, R. E. (2015). Pubertal testosterone influences threat-related amygdala- orbitofrontal cortex coupling. Social Cognitive and Affective Neuroscience, 10(3), 408-415.
doi: 10.1093/scan/nsu062 pmid: 24795438 |
| [66] |
Spinhoven, P., de Rooij, M., Heiser, W., Smit, J. H., & Penninx, B. W. (2012). Personality and changes in comorbidity patterns among anxiety and depressive disorders. Journal of Abnormal Psychology, 121(4), 874-884.
doi: 10.1037/a0028234 pmid: 22564181 |
| [67] |
Sprengelmeyer, R., Steele, J. D., Mwangi, B., Kumar, P., Christmas, D., Milders, M., & Matthews, K. (2011). The insular cortex and the neuroanatomy of major depression. Journal of Affective Disorders, 133(1-2), 120-127.
doi: 10.1016/j.jad.2011.04.004 pmid: 21531027 |
| [68] |
Sridharan, D., Levitin, D. J., & Menon, V. (2008). A critical role for the right fronto-insular cortex in switching between central-executive and default-mode networks. Proceedings of the National Academy of Sciences, 105(34), 12569-12574.
doi: 10.1073/pnas.0800005105 URL |
| [69] |
Surguladze, S. A., El-Hage, W., Dalgleish, T., Radua, J., Gohier, B., & Phillips, M. L. (2010). Depression is associated with increased sensitivity to signals of disgust: A functional magnetic resonance imaging study. Journal of Psychiatric Research, 44(14), 894-902.
doi: 10.1016/j.jpsychires.2010.02.010 pmid: 20307892 |
| [70] |
Suslow, T., Konrad, C., Kugel, H., Rumstadt, D., Zwitserlood, P., Schöning, S., ... Dannlowski, U. (2010). Automatic mood- congruent amygdala responses to masked facial expressions in major depression. Biological Psychiatry, 67(2), 155-160.
doi: 10.1016/j.biopsych.2009.07.023 URL |
| [71] |
Tang, S., Lu, L., Zhang, L., Hu, X., Bu, X., Li, H., ... Huang, X. (2018). Abnormal amygdala resting-state functional connectivity in adults and adolescents with major depressive disorder: A comparative meta-analysis. EBioMedicine, 36, 436-445.
doi: S2352-3964(18)30368-2 pmid: 30316866 |
| [72] |
Wardenaar, K. J., Conradi, H. J., Bos, E. H., & de Jonge, P. (2014). Personality modulates the efficacy of treatment in patients with major depressive disorder. The Journal of Clinical Psychiatry, 75(9), e916-923.
doi: 10.4088/JCP.13m08855 URL |
| [73] | Wang, J., Wang, N. N., & Zhang, Y. (Eds). (2013). Students' Mental Health. Beijing Normal University Press. |
| [王洁, 王宁宁, 张艳. (主编). (2013). 大学生心理健康. 北京: 北京师范大学出版社.] | |
| [74] | Yan, C., & Zang, Y. (2010). DPARSF: A MATLAB toolbox for "pipeline" data analysis of resting-state fMRI. Frontiers in Systems Neuroscience, 4, 13. |
| [75] | Yan, R., Geng, J. T., Huang, Y. H., Zou, H. W., Wang, X. M., Xia, Y., ... Lu, Q. (2022). Aberrant functional connectivity in insular subregions in somatic depression: A resting-state fMRI study. BMC Psychiatry, 22(1), 146. |
| [76] |
Yin, K., Zhao, J., Zhou, J., & Nie, Q. (2021). The big-five personality profiles: A person-centered approach. Advances in Psychological Science, 29(10), 1866-1877.
doi: 10.3724/SP.J.1042.2021.01866 |
|
[尹奎, 赵景, 周静, 聂琦. (2021). “大五”人格剖面: 以个体为中心的研究路径. 心理科学进展, 29(10), 1866-1877.]
doi: 10.3724/SP.J.1042.2021.01866 |
|
| [77] | Yoon, K. L., Maltby, J., & Joormann, J. (2013). A pathway from neuroticism to depression: Examining the role of emotion regulation. Anxiety, Stress & Coping, 26(5), 558-572. |
| [78] |
Zhang, T., Bai, T., Xie, W., Wei, Q., Lv, H., Wang, A., ... Wang, K. (2021). Abnormal connectivity of anterior-insular subdivisions and relationship with somatic symptom in depressive patients. Brain Imaging and Behavior, 15(4), 1760-1768.
doi: 10.1007/s11682-020-00371-x |
| [79] |
Zuroff, D. C., Mongrain, M., & Santor, D. A. (2004). Conceptualizing and measuring personality vulnerability to depression: Comment on Coyne and Whiffen (1995). Psychological Bulletin, 130(3), 489-511.
pmid: 15122935 |
| [1] | 刘德鹏, 李珏兴, 张生军, 庞旭宏, 王政. 为家所困还是弥补缺憾?负面家庭事件对有效领导行为的影响[J]. 心理学报, 2024, 56(5): 612-629. |
| [2] | 李为, 边子茗, 陈曦梅, 王俊杰, 罗一君, 刘永, 宋诗情, 高笑, 陈红. 9~12岁儿童应激与额颞区的关联: 来自多模态脑影像的证据[J]. 心理学报, 2023, 55(4): 572-587. |
| [3] | 徐健捷, 张一一, 林德堃, 车俐颖, 宋漫漫, 韩卓. 亲子依恋与儿童抑郁症状的关系:儿童对环境的生物敏感性的作用及父母差异[J]. 心理学报, 2023, 55(3): 469-480. |
| [4] | 郝子雨, 李欢欢, 林亦轩. 抑郁症自杀未遂者的痛苦逃避与背外侧前额叶−脑岛有效连接特征[J]. 心理学报, 2023, 55(12): 1966-1978. |
| [5] | 蔡惠燕, 苗心, 王鹏飞, 林志为, 王孟成, 杨文登, 麻彦坤, 曾红. 长期戒断海洛因成瘾者冲动性相关脑区的结构及功能特征[J]. 心理学报, 2021, 53(8): 861-874. |
| [6] | 莫李澄, 郭田友, 张岳瑶, 徐锋, 张丹丹. 激活右腹外侧前额叶提高抑郁症患者对社会疼痛的情绪调节能力:一项TMS研究[J]. 心理学报, 2021, 53(5): 494-504. |
| [7] | 李依蔓, 刘程, 庄恺祥, 霍腾宾, 徐鹏飞, 罗跃嘉, 邱江. 人格特质及脑功能连接对社交网络的影响[J]. 心理学报, 2021, 53(12): 1335-1347. |
| [8] | 陈玉明, 李思瑾, 郭田友, 谢慧, 徐锋, 张丹丹. 背外侧前额叶对主动遗忘负性社会反馈的作用:针对抑郁症的TMS研究[J]. 心理学报, 2021, 53(10): 1094-1104. |
| [9] | 罗扬眉, 莫凡, 陈煦海, 蒋宏达, 游旭群. 快乐型和实现型情感的习惯化 *[J]. 心理学报, 2018, 50(9): 985-996. |
| [10] | 范兴华, 方晓义, 黄月胜, 陈锋菊, 余思. 父母关爱对农村留守儿童抑郁的影响机制:追踪研究 *[J]. 心理学报, 2018, 50(9): 1029-1040. |
| [11] | 任志洪; 李献云; 赵陵波; 余香莲; 李政汉; 赖丽足; 阮怡君; 江光荣. 抑郁症网络化自助干预的效果及作用机制 ——以汉化MoodGYM为例[J]. 心理学报, 2016, 48(7): 818-832. |
| [12] | 陈乐妮; 王桢; 骆南峰;罗正学. 领导−下属外向性人格匹配性与下属工作投入的关系:基于支配补偿理论[J]. 心理学报, 2016, 48(6): 710-721. |
| [13] | 罗利;黄敏儿. 情绪调节在特质与情绪间中介效应模型的年龄差异[J]. 心理学报, 2016, 48(11): 1455-1466. |
| [14] | 刘小禹;刘军;许浚;吴蓉蓉. 职场排斥对员工主动性行为的影响机制 ——基于自我验证理论的视角[J]. 心理学报, 2015, 47(6): 826-836. |
| [15] | 吴梦莹;周仁来;黄雅梅;王庆国;赵燕;刘雁峰. 神经质程度和月经周期对女性主观情绪和生理反应的影响[J]. 心理学报, 2014, 46(1): 58-68. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||