

1 引言
创造性思维是个体以新的方式解决问题的高级思维活动(Sternberg & Lubart, 1996)。有研究表明, 执行功能与创造性思维相关(Beaty, Silvia, Nusbaum, Jauk, & Benedek, 2014; Benedek et al., 2014a; Benedek, Jauk, Sommer, Arendasy, & Neubauer, 2014b)。执行功能是控制个体思想和行为的基本认知加工能力, 与前额叶皮层相关, 由转换、抑制和工作记忆刷新三个成分组成(周雅, 2013)。近10多年来, 认知抑制与创造性思维的关系成为研究热点之一(Benedek, Franz, Heene, & Neubauer, 2012; Radel, Davranche, Fournier, & Dietrich, 2015; Zhang et al., 2016)。认知抑制是对认知内容或过程的抑制, 其在创造性思维中扮演着重要角色(Harnishfeger, 1995; 白学军, 巩彦斌, 胡卫平, 韩琴, 姚海娟, 2014; 胡卫平, 程丽芳, 贾小娟, 韩蒙, 陈英和, 2015; 张克, 杜秀敏, 仝宇光, 2017)。
对于创造性思维与认知抑制的关系, 研究者提出不同理论加以解释。首先, 有研究者提出了创造性思维的认知去抑制假说(以下简称认知去抑制假说) (Eysenck, 1995)。该假说认为, 相比低创造性思维水平者(以下称低创者), 高创造性思维水平者(以下称高创者)的多巴胺分泌水平升高, 血清素降低, 导致认知抑制能力降低, 并且其采用稳定的离焦注意的模式, 能够更多注意并记忆无关信息(Vartanian, 2002), 更具冲动性(Burch, Hemsley, Pavelis, & Corr, 2006), 潜在抑制能力缺乏(Peterson & Carson, 2000; Peterson, Smith, & Carson, 2002; Carson, Peterson, & Higgins, 2003; Chirila & Feldman, 2012)。研究发现, 低抑制有助于创造性观点产生(Radel, et al., 2015; Zhang et al., 2016)。通过降低大脑左额叶的活动并增加大脑右额叶的活动来降低认知抑制, 也发现减少的认知抑制对创造性观点产生有积极的影响(Mayseless & Shamay-Tsoory, 2015)。
随后, 有研究者提出了创造性思维的认知抑制假说(以下简称认知抑制假说) (Groborz & Necka, 2003)。该假说认为总体而言, 高创者的认知抑制能力高于低创者。以往研究采用Stroop 任务发现, 高创者在字色不一致条件下的反应时显著短于低创者(Edl, Benedek, Papousek, Weiss, & Fink, 2014; Groborz & Necka, 2003)。其他研究采用随机动作生成测验测量个体抑制能力, 结果也发现认知抑制与观点的流畅性、自我报告的创造性行为以及创造性成就呈正相关(Benedek et al., 2012; Zabelina, Robinson, Council, & Bresin, 2012)。执行功能与创造力关系的研究表明, 高执行功能导致高创造力。执行功能子成分中的抑制和更新可以预测创造力(Benedek et al., 2014b)。神经影像学研究也发现前额皮层的过度活化有益于产生创造性成果, 表明良好的认知控制促使更具创造性的观点产生(Colombo, Bartesaghi, Simonelli, & Antonietti, 2015)。
上述有关认知抑制与创造性思维的两种假说中, 前一种假说强调创造性思维的自动化加工, 认为低认知抑制在创造性加工中起重要作用; 后一种假说则强调创造性加工过程中控制性加工的重要性, 认为高水平的创造力需要一定程度的集中的心理努力, 高认知抑制有助于创造性问题的发现和解决(Groborz & Nęcka, 2003)。然而, 创造性加工过程中可能既有自动化加工, 也有控制性加工, 只是在不同的阶段或不同的情境下个体更倾向于采用某种加工模式。Kris (1952)提出创造性思维的初级-次级加工理论, 初级加工具有自我中心、自由联想、无拘束的特点, 次级加工常常和有意识、有目的的思考相联系, 高创者更擅于在这两种加工模式之间进行转换。因此, 高创者的认知去抑制和认知抑制两种状态均可以运作, 认知去抑制可能是其面对模糊问题或处于某个阶段而表现出来的特质(Cheng, Hu, Jia, & Runco, 2016; Zabelina & Beeman, 2013; Radel et al., 2015; 姚海娟, 白学军, 2014)。
近来, 有研究者对创造性思维的认知抑制和认知去抑制假说进行整合, 提出创造性思维的适应性认知抑制假说(以下简称适应性认知抑制假说) (Martindale, 1999, 2007)。该假说认为, 高创者面对不同任务要求时会对注意焦点进行灵活转换, 表现出较高的目标导向的注意技能, 相比低创者, 高创者能够更好地根据任务要求调整注意的焦点。在创造性问题解决的早期阶段, 问题相对地定义不良, 高创者更可能倾向于离焦注意, 但注意宽泛且导致信息加工速度变慢; 在创造性问题解决的晚期阶段, 高创者从抑制无关信息和增加对任务的集中注意中获益, 注意变窄并提高了任务加工速度(Ansburg & Hill, 2003; Vartanian, 2009; Zabelina & Beeman, 2013), 并能够根据问题解决结果灵活调整问题解决策略(Zabelina & Robinson, 2010)。Cheng等人(2016)探讨了认知抑制在创造性的问题发现的早期和晚期阶段中的作用。结果发现, 个体的认知抑制与问题发现任务中思维的流畅性和灵活性有关, 但与独创性无关; 低认知抑制提高了一开始的独创性, 但在随后的加工中, 高认知抑制有助于创造性加工。这表明创造性的问题发现的早期和晚期阶段需要不同的认知抑制水平。张克等人(2017)采用定向遗忘范式, 探讨创造性思维水平高低与认知抑制的关系。结果发现, 低创者在2 s和5 s以及高创者在5 s时间间隔时, 均对中性词表现出定向遗忘效应, 而对负性词没有表现出定向遗忘效应。高创者在2 s时间间隔下, 对中性和负性词均表现出定向遗忘效应。结果表明较短时间内高创者对负性情绪的认知抑制能力优于低创者。还有研究发现, 创造性思维水平与无干扰任务(如概念判断任务)的反应时呈负相关, 与包含干扰的任务(如负启动任务)的反应时呈正相关(Dorfman, Martindale, Gassimova, & Vartanian, 2008; Vartanian, Martindale, & Kwiatkowski, 2007)。但是, 这两个研究采用了两种不同干扰水平的任务, 没有单独从认知抑制任务来探讨高创者的适应性认知抑制。因此, 如果高创者的认知抑制具有适应性, 那么这种认知抑制能力是更高更灵活, 还是更低更灵活呢?这一问题还需要采用单独测定认知抑制的任务范式进行检验。
前人研究发现, Stroop任务是测量认知抑制的有效研究范式(Edl et al., 2014; Groborz & Necka, 2003; Zabelina & Robinson, 2010)。该任务要求个体判断字义或者判断字色。字色不一致条件下的反应时通常比字色一致时的反应时要长(Stroop, 1935)。所以本研究的第一个目的是采用经典Stroop任务来考察认知抑制与创造性思维的关系, 检验认知去抑制和认知抑制两种假说。在实验1中, 我们采用经典Stroop任务, 如果高创者的反应时干扰效应量显著短于低创者, 则表明高创者的认知抑制能力较强, 相反则表明高创者的认知抑制能力较弱。
但是, 实验1仅能对认知去抑制和认知抑制两个假说进行验证, 无法检验适应性认知抑制假说。基于适应性认知抑制假说的观点, 我们预期任务因素可能调节认知抑制与创造性思维的关系。研究发现, 时间压力对创造性思维有影响, 但结论并不一致。一种观点认为, 时间压力降低个体的创造性。例如, Amabile, Hadley和Kramer (2002)调查了177名从事需要高度创意工作的项目人员的9000多篇日记, 发现他们在需要争分夺秒地完成工作任务时表现出的创意最差; 另一种观点认为, 时间压力促进个体的创造性(Darini, Pazhouhesh, & Moshiri, 2011), 或者时间压力能够促进思维的流畅性(张景焕, 王亚男, 初玉霞, 徐希铮, 2011)。本研究采用Szollos (2009)对时间压力的界定, 即个体对没有足够时间做事情的一种认知体验。所以, 本研究的第二个目的是探讨时间压力对认知抑制与创造性思维的调节作用。在实验2中, 采用更灵活的Stroop范式, 操纵有和无时间压力两种任务情境, 验证适应性认知抑制假说。从信息加工的角度分析, 时间压力会影响个体在任务环境中的注意对象和范围, 个体会更关注与任务相关的特征, 而忽略与之不相关的特征, 因此预期高创者比低创者在抑制冲突刺激时更能从时间压力情境下获益。
研究表明, 个体的认知加工与自主唤醒有关(Gendolla, Wright, & Richter, 2012)。Pennebaker和Chew (1985)提出, 抑制需要努力, 反过来又导致生理成本的增加。因此, 实验2拟增加生理指标来反映高低创者在不同时间压力任务情境下个体内部的唤醒情况。目前采用皮肤电活动(electrodermal activity, EDA)来评定自主神经系统的动力学特点已经非常普及(Bach & Friston, 2013)。自主神经系统是身体内部的调控网络, 通过平衡交感神经系统和副交感神经系统来反映个体感知到的压力水平。EDA是一种测量皮肤电导变化的方法, 具有无创性, 反映交感神经系统的激活水平(Posada-Quintero et al., 2016)。许多研究采用皮肤电反应(skin conductance responses, SCRs)这一指标, 因为SCRs表示的是EDA信号的快速时相转移(Boucsein et al., 2012), 能够较快地反映呈现的刺激所引起的皮肤电活动的变化。以往研究表明, Stroop任务中的集中注意、冲突探测以及错误意识与补偿等加工与自主唤醒有关(Kobayashi, Yoshino, Takahashi, & Nomura, 2007)。相比字色一致条件或中性条件, 字色不一致条件会引起更大的皮肤电活动变化(Kobayashi et al., 2007; Lemche et al., 2016), 表明个体抑制冲突并解决会引起交感神经系统更高强度的激活。抑制与生理唤醒的间接支持证据来自于情绪反应调节的研究。研究表明, 个体抑制自身的情绪反应会产生交感神经兴奋, 导致皮肤电活动变化增加, 手指脉冲幅度增加, 指温升高(Demaree et al., 2006)。同样, 犯罪测谎的研究表明, 犯罪者在测谎时对抗条件下比合作条件下引起了更大的皮肤电活动变化(Zvi, Nachson, & Elaad, 2012)。研究者认为, 皮肤电活动变化的增强反映了个体有更高的防御动机并尝试抑制对抗条件下的生理唤醒。
上述表现只是个体内部的抑制增加会导致皮肤电生理唤醒的增加, 然而皮肤电活动变化还可能反映无意识的评价过程, 是对相关的即将发生的刺激的可能(消极)结果或后果的预期, 也可能反映无意识的情感加工, 可以作为指导个体未来决策的一个身体标记, 帮助个体在有风险或不确定的情况下指引方向并采取行动(Dawson, Schell, & Courtney, 2011)。对于高创者和低创者来说, 他们可能具有不同的生理表现(Kwiatkowski, 2002; Martindale, 1999; 谷传华等, 2015)。相比低创者, 高创者往往有较高的基础唤醒水平, 表现为较低的α波活动和较高的皮肤电导水平(Martindale, 1999), 但其在不需要创造性的任务(如爱荷华赌博任务)中的皮肤电活动变化较低(Galang, Castelo, Santos, Perlas, & Angeles, 2016), 在需要发散思维的创造任务中交感激活水平更高(Silvia, Beaty, Nusbaum, Eddington, & Kwapil, 2014)。我们预期, 如果符合适应性认知抑制假说, 则高创者在不同任务条件下会表现出变化的生理唤醒水平, 而低创者则会表现出固定的生理唤醒水平。如果高创者比低创者的认知抑制能力强, 那么相比无时间压力情境, 高创者在有时间压力情境下可能不会表现出皮肤电活动变化的显著差异, 而低创者在更有时间压力的任务情境下较难根据任务要求完成较快反应, 因此需要更大的抑制努力, 其皮肤电活动变化会显著增加。
综上所述, 本研究拟在以往研究的基础上, 采用Stroop颜色命名任务和更具有灵活性的Stroop字义-颜色命名转换任务和操纵时间压力, 探讨高低创造性思维水平者认知抑制能力的差异。我们预测, 如果高创者比低创者的认知抑制能力更高更灵活, 则从行为指标上, 高创者的Stroop干扰效应量显著小于低创者, 高创者在有时间压力条件下的干扰效应量显著小于无时间压力条件下, 而低创者则没有; 从皮肤电指标上, 高创者在不同条件下的生理唤醒水平不同, 而低创者则基本保持一致的生理唤醒水平。
2 实验1:高低创造性思维水平者的认知抑制能力比较
2.1 方法
2.1.1 被试
对568名大学生施测《托兰斯创造性思维测验》(TTCT)词汇卷和图画卷, 删除没有完成测试的被试15人, 最终有效问卷553份, 其中男生239人, 女生314人, 平均年龄19.74 ± 1.06岁。
将各维度得分转换为标准分后求总分, 根据总分排序, 并自愿参加的原则, 选取高创者30人(男13人, 女17人), 平均年龄19.60岁, 低创者30人(男12人, 女18人), 平均年龄19.55岁。高创组(M = 325.87, SD = 21.67)和低创组(M = 108.50, SD = 23.49) 的TTCT得分进行独立样本t检验的差异显著, t(58) = 37.25, p < 0.001。采用《瑞文标准推理测验》测量被试的智力, 对高创组(M = 52.03, SD = 3.89)和低创组(M = 49.83, SD = 4.79)的智力得分进行独立样本t检验的差异不显著, t(58) = 1.95, p > 0.05。被试无智力障碍, 无色盲、色弱, 实验完成后给每个被试适量报酬。
2.1.2 实验设计
实验设计为2(创造性组别:高创组、低创组)×2(刺激类型:字色一致、字色不一致)两因素混合设计。其中创造性组别为组间变量, 刺激类型为组内变量。因变量为反应时和正确率。
2.1.3 仪器和材料
实验仪器使用美国DELL笔记本电脑, 实验材料通过13.3英寸液晶显示器呈现。分辨率为1024× 768。
采用《托兰斯创造性思维测验》(TTCT)中文修订版(叶仁敏, 洪德厚, 保尔•托兰斯, 1988)的词汇卷和图画卷对被试的创造性思维水平进行测量。评分方法:本测验由两名心理学专业本科生(1男1女)独立进行评分。经过对二人的评分结果进行相关分析, 结果表明, 词汇卷的评分者一致性为0.90, p < 0.001; 图画卷的评分者一致性为0.89, p < 0.001, 说明二人评分信度较高, 测验成绩取二者的平均值。
选用张厚粲和王晓平 (1989)修订的《瑞文标准推理测验》中国城市修订版。测验共60道题目。测验限时40分钟, 分半信度为0.95, 间隔半个月的重测信度为0.82, 间隔一个月的重测信度为0.79, 同时效度为0.71, 具有良好的信效度。
采用经典的Stroop任务, 刺激材料是“红”和“绿”两个汉字, 分别用“红”和“绿”两种颜色书写。每个字的大小是1.1°×1.1°, 形成字色一致刺激2种, 字色不一致刺激2种, 背景为黑色。刺激总数共有96个, 其中字色一致的刺激48个, 不一致刺激48个。练习材料为红色和绿色方块, 练习20次。实验中所使用的图片文件都是由Photoshop 7画图软件制作而成。实验程序采用E-prime 1.1编制。
2.1.4 实验程序
正式实验之前先分批对被试进行创造性思维测验和智力测验, 每批大约20~40人。给被试施测《托兰斯创造性思维测验》的词汇卷和图画卷, 以及《瑞文标准推理测验》。词汇卷、图画卷和智力测验之间皆让被试休息15分钟。挑选的符合条件的被试再通过电话预约来实验室做实验。
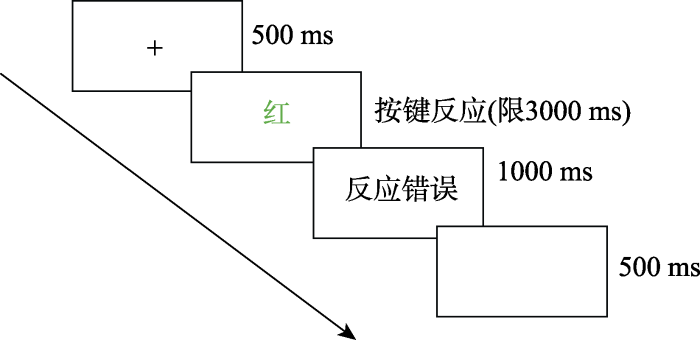
正式实验在实验室内完成。实验室安静舒适, 隔音较好。被试坐在距离计算机屏幕约60 cm的位置。整个实验分练习和正式实验, 两个阶段实验程序一致。实验程序如图1所示。首先在屏幕中央出现一个注视点“+”500 ms, 然后出现一个实验刺激(颜色字), 程序允许的反应时间为3000 ms, 被试正确反应之后与下一个实验刺激之间的时间间隔为500 ms。被试错误反应之后在屏幕上呈现1000 ms的反馈错误信息作为惩罚, 目的是保证高正确率, 然后分析反应时, 正确反应后无反馈信息。要求被试对出现的刺激字的颜色又快又准地按键作出反应。实验时间为5 min。
图1
2.2 结果
数据采用SPSS 18.0进行分析。删除反应时超过2000 ms的数据, 并剔除超过平均数3个标准差之外的极端数据。删除反应错误的数据占总数据的3.72%。删除的数据占总数据的4.80%。对高创组和低创组被试在Stroop颜色命名任务上字色一致条件和字色不一致条件下的正确反应时和正确率进行分析。
对反应时进行了2(创造性组别:高创组、低创组)×2(刺激类型:字色一致、字色不一致)的重复测量方差分析, 结果发现(见图2):创造性组别的主效应显著, F(1, 58) = 8.46, p < 0.01, ηp2 = 0.13, 高创者的反应时显著短于低创者; 刺激类型的主效应显著, F(1, 58) = 24.92, p < 0.001, ηp2 = 0.30, 字色一致条件下的反应时显著小于字色不一致条件下的反应时; 创造性组别和刺激类型的交互作用显著, F(1, 58) = 9.26, p < 0.05, ηp2 = 0.14。进一步分析发现, 高创者在字色一致条件上的反应时显著短于低创者, p < 0.05, 且高创者在字色不一致条件下的反应时也显著小于低创者, p < 0.01。将被试在字色不一致条件下的反应时减去字色一致条件下的反应时得出反应时的干扰效应量, 然后对高创组和低创组的反应时干扰效应量进行独立样本t检验, 结果发现, 两组的反应时干扰效应量存在显著差异, t(58) = -3.04, p < 0.01, 高创者的干扰效应量(M = 7 ms, SD = 25 ms)显著小于低创者(M = 27 ms, SD = 27 ms)。
图2

对正确率数据(见表1)的分析发现:刺激类型的主效应显著, F(1, 58) = 11.33, p < 0.01, ηp2 = 0.16, 字色一致条件下的反应正确率显著高于字色不一致条件; 创造性组别和刺激类型的交互作用显著, F(1, 58) = 4.74, p < 0.05, ηp2 = 0.08, 进一步分析发现, 高创者在字色一致和不一致条件下的正确率差异不显著, p > 0.05, 而低创者在字色不一致条件下的正确率显著低于字色一致条件, p < 0.01。将被试在字色一致条件下的正确率减去字色不一致条件下的正确率得出正确率的干扰效应量, 然后对高创组和低创组的正确率干扰效应量进行独立样本t检验, 结果发现, 两组的正确率干扰效应量存在显著差异, t(58) = -2.18, p < 0.05, 高创者的干扰效应量(M = 0.63%, SD = 3.88%)显著小于低创者(M = 2.92%, SD = 4.26%)。创造性组别的主效应不显著, F(1, 58) < 1。
表1 高创组和低创组在各实验条件下的正确率(%) [M (SD)]
| 创造性组别 | 字色一致 | 字色不一致 |
|---|---|---|
| 高创组 | 97.60(3.55) | 96.98 (2.39) |
| 低创组 | 98.64(2.55) | 95.73(4.14) |
2.3 讨论
从实验1的反应时和正确率结果来看, 相比低创者, 高创者的反应时干扰效应量和正确率干扰效应量更小。这与前人的研究结果是一致的(Benedek et al., 2014b; Edl et al., 2014; Zmigrod, Zmigrod, & Hommel, 2015)。通过实验1检验了创造性思维的认知去抑制和认知抑制这两个假说, 结果支持了创造性思维的认知抑制假说, 表明高创者的认知抑制能力强于低创者。然而, 实验1的结果与Groborz和Nęcka (2003)的研究结果并不完全一致。Groborz和Nęcka (2003)的研究仅发现, 高创者在不一致条件下的反应更有规律。这可能与两个研究的实验材料和实验程序不同有关。相比之下, 本研究中实验1所采用的实验材料和实验程序更简单, Stroop任务的刺激集为2, 仅红、绿两种颜色字, 且仅要求被试对字的颜色进行命名。但是, Groborz和Nęcka (2003)研究中Stroop任务的刺激集为5, 而且被试要依据屏幕下方的提示线索来确定进行颜色命名还是字义命名, 研究程序更有难度。因此, 仅采用简单的Stroop任务来揭示认知抑制与创造性思维的关系并不全面。
所以, 实验2拟增大刺激集大小, 采用更灵活的Stroop字义-颜色命名转换任务, 颜色字包括红、绿、黄三种, 并操纵有和无时间压力条件, 进一步检验创造性思维的适应性认知抑制假说。反应时指标仅能反映个体的外部反应, 因此实验2拟增加皮肤电生理指标以反映不同创造性思维水平者抑制过程的内部生理唤醒差异。
3 实验2:时间压力对高低创造性思维水平者认知抑制的影响
3.1 方法
3.1.1 被试
从实验1施测TTCT的总被试中, 根据TTCT总分排序, 挑选没有参加实验1的高创者和低创者各26人。因为4名被试的皮肤电线显示干扰很大且无起伏被删除。最终被试48名。高创组24人(男11人, 女13人), 平均年龄19.33岁; 低创组24人(男12人, 女12人), 平均年龄19.21岁。高创组(M = 301.43, SD = 50.71)和低创组(M = 130.17, SD = 24.64) 的TTCT得分进行独立样本t检验的差异显著, t(46) = 14.88, p < 0.001。采用《瑞文标准智力测验》测量被试的智力, 高创组(M = 54.17, SD = 3.52)和低创组(M = 51.58, SD = 5.57)的智力得分进行独立样本t检验的差异不显著, t(46) = 1.92, p > 0.05。被试无智力障碍, 无色盲、色弱, 实验完成后给每个被试适量报酬。
3.1.2 实验设计
2(创造性组别:高创组、低创组)×2(时间压力情境:有、无)×2(任务类型:字义命名、颜色命名)×2(刺激类型:一致、不一致)四因素混合设计, 其中创造性组别和时间压力情境为组间变量, 任务类型和刺激类型为组内变量。
3.1.3 仪器和材料
实验采用Superlab 4.5刺激呈现系统呈现刺激并记录被试的反应, 该系统刺激呈现与计时精度均为1 ms。刺激通过DELL 23英寸显示器呈现, 被试距屏幕1 m处。显示器的分辨率为1024×768。屏幕的背景为黑色。
使用BIOPAC MP150型16导无线生理记录仪系统的信号探测器、转换器和放大器等系统, 记录被试在静止阶段和实验阶段的皮肤电。具体参数为16个模拟通道输入, 2个模拟通道输出, 16个数字通道输入; 最大输入电压:±12 V; 输入阻抗:1 MΩ; A/D转换率:16 Bits。本研究中皮肤电值的单位是毫西门子(microsimens)。
实验材料是“蓝”、“红”、“绿”色印刷的“蓝”、“红”和“绿”三个字。每个字的大小是1.1°×1.1°。共96个试次, 其中一致条件48个, 不一致条件48个, 分为4个block, 每个block包括24个, 其中一半为一致刺激, 一半为不一致刺激。第一个与第三个Block要求被试对刺激的字义进行按键反应, 第二个与第四个Block要求被试对刺激的颜色进行按键反应。练习任务材料为24个颜色词刺激, 要求对字义进行命名。
3.1.4 实验程序
实验采用个别施测, 具体的实验程序如下:
第一步:被试坐在被试座椅上, 与电脑显示屏距离约为1 m, 主试向其简单介绍需要进行的实验任务, 并给被试连接上记录皮肤电反应活动的传感器。
第二步:采集基线。要求被试心情平静下来, 头脑中什么也不要想, 尽量保持身体不动, 采集5 min的生理指标作为基线值。
第三步:给被试详细说明实验指导语后开始练习任务。练习任务结束后让被试对其目前所感受到的时间压力进行评定, “1”代表根本没有, “5”代表非常强烈。然后进行正式实验任务。
第四步:正式实验。随机分配高创者和低创者进入有或无时间压力实验条件。时间压力情境的设置主要考虑被试的反应时, 本实验中给予“反应太慢”试验的反应时临界值的界定主要是参考Kobayashi等(2007)的文献中测量的各条件下的反应时平均数, 最后将该值定为550 ms。
被试先做字义命名任务, 然后按顺序依次为颜色命名任务、字义命名任务和颜色命名任务。不同时间压力情境下的具体实验程序见图3所示。
图3
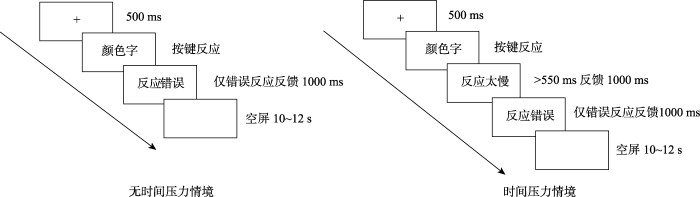
有时间压力情境下:首先在屏幕中央出现一个注视点“+”500 ms, 然后出现一个实验刺激(颜色字), 程序允许被试的反应时间为3000 ms。当被试的反应超过550 ms, 则在屏幕上给被试呈现反馈信息“反应太慢”, 如果被试反应错误, 则在屏幕上给被试呈现反馈信息“反应错误”, 被试正确反应之后与下一次trial之间的时间间隔为10~12 s。字义命名任务要求被试忽略字的颜色, 对屏幕上出现的字的字义进行又快又准的按键反应。被试通过外接反应盒进行反应。用右手的食指、中指和无名指在反应盒上进行按键反应, 按键上有相对应的颜色, 绿字按第一个键, 红字按第二个键, 蓝字按第三个键。颜色命名任务则要求被试对字的颜色进行又快又准的按键反应。
无时间压力情境下:没有“反应太慢”的反馈, 其它同上。
第五步:所有被试在实验结束后立即对感受到的完成任务的时间压力进行评定, “1”代表根本没有, “5”代表非常强烈。
3.1.5 生理数据的采集与分析
皮肤电采集:对被试的左手食指和中指的末端指腹用酒精消毒后, 贴上电极片, 然后将电极线的一端连接在电极片上, 电极线的另一端插在无线发射器测量皮肤电反应的EDA端口上, 信号发射到生理记录仪的GSR100C模块上记录被试的皮肤电, 采样率为1000 Hz。
参照Kobayashi等人(2007)的研究, 取刺激呈现之后1~4 s探测到的皮肤电活动的最大值作为该trial的皮肤电反应值。各条件下皮肤电活动变化为皮肤电反应最大值减去基线值。为了减少数据分布的偏度, 将皮肤电值+1之后进行对数转换再进行统计分析。
3.2 结果
统计反应时数据时, 删除反应时大于2000 ms的极端值, 剔除超过平均数3个标准差之外的极端数据, 删除错误反应的数据8.31%。删除数据占总数据的10.33%。
所采集的生理数据在Acqknowledge 4.2软件上进行编辑处理。参照Kobayashi等人(2007)的研究, 将SCR的最小起伏值定为0.05 microsimens, 将不满足这一标准的数据删除。删除的数据占总数据的7.82%。后期数据采用SPSS 18.0进行分析。
对被试是否感受到时间压力的主观评定进行独立样本t检验, 结果发现, 两组被试完成实验后的主观评定结果差异显著, t(46) = 14.86, p < 0.001, 有时间压力组(M = 3.83, SD = 0.48)感受到的时间压力显著高于无时间压力组(M = 1.21, SD = 0.51)。结果表明, 实验中对被试设置的时间压力情境程序确实引起了被试较强的时间压力感受。
3.2.1 高创组和低创组在不同时间压力情境下的反应时
根据高创组和低创组被试在Stroop字义命名和颜色命名任务上不同时间压力条件下的反应时进行统计, 结果如图4所示。
图4

对反应时进行2(创造性组别:高创组、低创组)×2(时间压力情境:有、无)×2(任务类型:字义命名、颜色命名)×2(刺激类型:一致、不一致)的重复测量方差分析, 结果发现:任务类型的主效应显著, F(1, 44) = 4.56, p < 0.05, ηp2 = 0.09, 字义命名任务的反应时显著短于颜色命名任务; 刺激类型的主效应显著, F(1, 44) = 127.82, p < 0.001, ηp2 = 0.74, 一致条件下的反应时显著短于不一致条件下的反应时; 时间压力情境的主效应显著, F(1, 44) = 34.12, p < 0.001, ηp2 = 0.44, 有时间压力情境下的反应时显著短于无时间压力情境下的反应时; 创造性组别的主效应不显著, F(1, 44) = 2.12, p > 0.05;
根据实验目的, 重点分析与创造性组别变量之间的显著的交互作用。结果发现, 刺激类型、时间压力情境与创造性组别三者的交互作用显著, F(1, 44) = 6.31, p < 0.05, ηp2 = 0.13。进一步分析发现, 对于高创者而言, 刺激类型的主效应显著, F(1, 23) = 70.59, p < 0.001, ηp2 = 0.75, 时间压力情境的主效应显著, F(1, 23) = 22.99, p < 0.001, ηp2 = 0.50, 时间压力情境和刺激类型的交互作用显著, F(1, 23) = 25.19, p < 0.001, ηp2 = 0.52, 对高创者在有和无时间压力情境下的干扰效应量进行独立样本t检验, t(23) = -5.02, p < 0.001, 即高创者在有时间压力情境下的干扰效应量(50 ms)显著短于无时间压力情境下(197 ms); 对于低创者而言, 刺激类型的主效应显著, F(1, 21) = 101.86, p < 0.001, ηp2 = 0.83, 时间压力情境的主效应显著, F(1, 21) = 19.23, p < 0.001, ηp2 = 0.48, 时间压力情境和刺激类型的交互作用不显著, F(1, 21) = 1.24, p > 0.05, ηp2 = 0.06, 对低创者在有和无时间压力情境下的干扰效应量进行独立样本t检验, t(21) = -1.11, p > 0.05, 即低创者在有时间压力条件下的干扰效应量(133 ms)和无时间压力条件下的干扰效应量(166 ms)无显著差异。
此外, 刺激类型与时间压力情境的交互作用显著, F(1, 44) = 15.44, p < 0.001, ηp2 = 0.26, 任务类型与刺激类型的交互作用显著, F(1, 44) = 10.92, p < 0.01, ηp2 = 0.20, 因与研究目的无关, 不作具体分析。刺激类型与创造性组别的交互作用不显著, F(1, 44) = 1.61, p > 0.05, 其他变量之间的交互作用均差异不显著, Fs (1, 44) < 1。
3.2.2 高创组和低创组在不同时间压力情境下的正确率
根据高创组和低创组被试在Stroop字义命名和颜色命名任务上的正确率进行统计, 结果如表2所示。
表2 高创组和低创组在不同时间压力情境下的正确率(%) [M (SD)]
| 时间压力 情境 | 创造性组别 | 字义命名 | 颜色命名 | ||
|---|---|---|---|---|---|
| 一致 | 不一致 | 一致 | 不一致 | ||
| 有时间压力 | 高创组 | 92.36 (7.29) | 82.29 (14.56) | 91.32 (6.52) | 83.33 (7.95) |
| 低创组 | 89.93 (6.76) | 80.21 (14.56) | 92.71 (7.13) | 80.21 (10.22) | |
| 无时间压力 | 高创组 | 96.88 (5.06) | 96.53 (3.91) | 99.31 (2.41) | 95.49 (5.46) |
| 低创组 | 98.61 (2.71) | 95.83 (4.35) | 98.96 (2.59) | 90.97 (7.07) | |
对正确率进行2(创造性组别:高创组、低创组)×2(时间压力情境:有、无)×2(任务类型:字义命名、颜色命名)×2(刺激类型:一致、不一致)的重复测量方差分析, 结果发现:刺激类型的主效应显著, F(1, 44) = 58.78, p < 0.001, ηp2 = 0.57, 一致条件下的反应正确率显著高于不一致条件; 时间压力情境的主效应显著, F(1, 44) = 36.12, p < 0.001, ηp2 = 0.45, 有时间压力条件下的反应正确率显著低于无时间压力条件; 任务类型的主效应不显著, F(1, 44) < 1; 创造性组别的主效应不显著, F(1, 44) < 1。
刺激类型与时间压力情境的交互作用显著, F(1, 44) = 12.39, p = 0.001, ηp2 = 0.22, 因与研究目的无关, 不作具体分析; 刺激类型与创造性组别的交互作用不显著, 任务类型与刺激类型的交互作用不显著, 任务类型、时间压力情境与创造性思维组别的交互作用不显著, 任务类型、刺激类型与时间压力情境的交互作用不显著, 任务类型、刺激类型与创造性思维组别的交互作用不显著, Fs (1, 44) > 1.03, ps > 0.10; 其他变量之间的交互作用均不显著, Fs (1, 44) < 1。
3.2.3 高创组和低创组在不同时间压力情境下不同条件的皮肤电结果
对被试的正确反应的皮肤电活动变化进行分析, 结果如图5所示。
图5
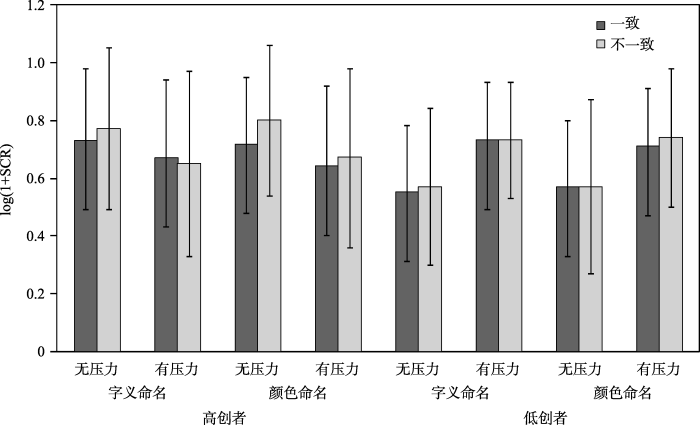
对皮肤电活动变化进行2(创造性组别:高创组、低创组)×2(时间压力情境:有、无)×2(任务类型:字义命名、颜色命名)×2(刺激类型:一致、不一致)的重复测量方差分析, 结果发现:任务类型的主效应不显著, F(1, 44) = 2.20, p > 0.05, ηp2 = 0.05; 刺激类型的主效应不显著, F(1, 44) < 1; 时间压力情境的主效应不显著, F(1, 44) < 1; 创造性组别的主效应不显著, F(1, 44) < 1;
根据实验目的, 重点分析与创造性组别变量之间的显著的交互作用。结果发现:时间压力情境与创造性组别的交互作用达到边缘显著, F(1, 44) = 3.14, p < 0.1 (p = 0.08), ηp2 = 0.07, 进一步分析发现, 高创者在无时间压力(0.75)和有时间压力(0.66)情境下的皮肤电活动变化无显著差异, p > 0.05, 低创者在无时间压力(0.57)和有时间压力(0.73)情境下的皮肤电活动变化也无显著差异, p > 0.05。
任务类型、刺激类型与创造性组别三者的交互作用达到边缘显著, F(1, 44) = 3.33, p < 0.1 (p = 0.08), ηp2 = 0.07; 进一步分析发现, 高创者在颜色命名任务中不一致条件下的皮肤电活动变化(0.74)显著高于一致条件(0.71), p < 0.05, 高创者在字义命名任务中一致条件和不一致条件下的皮肤电活动变化差异不显著, p > 0.05, 但是低创者在字义命名任务和颜色命名两个任务中一致和不一致条件下的皮肤电活动变化皆没有显著差异, p > 0.05。
此外, 任务类型与刺激类型的交互作用显著, F(1, 44) = 7.38, p < 0.01, ηp2 = 0.14, 因与研究目的无关, 不做具体分析; 刺激类型与时间压力情境的交互作用不显著, 任务类型、时间压力情境与创造性思维组别三者的交互作用显著, 任务类型、刺激类型与时间压力情境的交互作用不显著, Fs (1, 44) > 1.10, ps > 0.10; 其他变量之间的交互作用均差异不显著, Fs (1, 44) < 1。
3.3 讨论
实验2反应时结果表明, 高创者在有时间压力条件下的干扰效应量显著小于无时间压力条件下, 而低创者在有和无时间压力条件下的干扰效应量无显著差异。这表明相比低创者, 高创者具有更灵活的认知抑制能力。此结果与研究预期和前人研究中认为时间压力对创造性有促进作用的结果是一致的(Darini et al., 2011; 张景焕等, 2011)。时间压力促进了个体创造性思维的流畅性维度。面对时间压力情境, 高创者能够调节自己的注意更集中于任务特征, 抑制无关特征的干扰, 更能从时间压力情境中获益, 表现出比低创者更高的认知抑制能力。综合来看, 相比低创者, 高创者能够在有和无时间压力情境下灵活调整注意焦点, 表现出更灵活的认知抑制水平。Silvia等(2014)的研究发现, 高创造性成就者从基线期到完成发散性思维任务期间交感神经活动显著增加, 并且他们能给出更具创造性的观点。研究者认为, 这反映高创造性成就者完成创造性任务时付出了更高的努力, 支持创造力的执行控制观点。
皮肤电指标结果发现, 高创者在颜色命名任务中不一致条件下的皮肤电活动变化显著高于一致条件, 在字义命名任务中无此差异, 低创者在字义和颜色命名两个任务各条件下的皮肤电活动变化均无显著差异。这表明, 高创者对不同任务表现出了不同的自主生理唤醒水平, 而低创者则表现出基本一致的自主生理唤醒水平。这与其他研究中高创者有面对不同任务表现出变化的生理特点的结果是一致的(Kwiatkowski, 2002; Martindale, 1999)。实验2结果表明, 相比低创者, 高创者在面对不同任务情境具有灵活地认知抑制水平和唤醒水平。
4 总讨论
本研究以Stroop颜色命名任务和Stroop字义-颜色命名转换任务作为测定认知抑制的范式, 考察了创造性思维与认知抑制的关系, 并探讨了时间压力在认知抑制与创造性思维关系中的作用。实验1结果表明, 高创者的认知抑制能力高于低创者; 实验2的反应时结果表明, 高创者在不同时间压力情境下能够灵活地调节自身的注意焦点, 而低创者则不能。实验2的皮肤电结果表明高创者面对不同的任务条件表现出变化的生理唤醒水平, 而低创者则表现出基本一致的生理唤醒水平。
如前言所述, 以往研究对创造性思维与认知抑制的关系一直存在争论(Benedek et al., 2012; Benedek et al., 2014b; Carson et al., 2003; Dorfman et al., 2008; Edl et al., 2014; Groborz & Nęcka, 2003; Vartanian et al., 2007; Zabelina et al., 2012)。从本研究的反应时结果来看, 实验1采用Stroop颜色命名任务发现, 相比低创者, 高创者的反应时干扰效应量和正确率干扰效应量更小。结果表明, 相比低创者, 高创者表现出更强的认知抑制能力, 也就是说高创者的一个认知特征是能够有效抑制优势的但是不相关的反应倾向(Edl et al., 2014)。这一结果与以往研究采用Stroop任务的研究结果一致(Benedek et al., 2014b; Edl et al., 2014), 也与采用随机动作产生任务(Benedek et al., 2012; Zabelina et al., 2012; 胡卫平等, 2015)以及遗忘效应任务(Storm & Patel, 2014; 张克等, 2017)的结果一致。这一结果还得到执行功能促进创造性思维的相关研究的支持(Jauk, Benedek, Dunst, & Neubauer, 2013; Jauk, Benedek, & Neubauer, 2014)。处理新颖性任务需要抑制性加工, 创造在很大程度上就等同于处理新任务和新情况, 创造过程需要对一些自动化反应的压抑, 从而产生一个新颖的、独创的、未预期的解决方法。此外, 在给定的情况下抵抗无关的一种反应趋势会促进独创性观点产生(Benedek et al., 2012; Benedek et al., 2014b)。
本研究结果与前人以整体-局部加工任务为认知抑制任务的研究结果是一致的(Groborz & Necka, 2003; Zmigrod et al., 2015)。本研究结果符合创造力的产生-评价理论对创造性行为的定义(Groborz & Necka, 2003)。根据该理论, 创造性行为不仅包括产生观点, 而且包括对产生独创性的产品的选择能力, 这种选择能力部分依赖于认知控制中的有效注意机制。此外, Zmigrod等(2015)的研究也发现, 发散思维中的灵活性可以显著预测整体-局部加工任务中的局部干扰效应(即识别刺激的整体特征受到来自局部水平信息的干扰), 发散思维的灵活性越高, 局部干扰效应越小。这表明如果个体更注意整体方面而没有被局部方面转移注意力, 那么个体更有可能找到针对某一个问题的各种各样的解决方案。局部干扰条件中, 被试在识别大字母的时候抑制来自小字母的干扰, 是一种抑制反应定势的倾向。这与Stroop任务中识别颜色字时需要抑制颜色与字义的冲突的干扰相似。
当前研究结果与认知去抑制假说冲突, 该假说认为认知去抑制可以提高产生许多不同想法的流畅性, 认知抑制水平低是高创造者的特征之一。Eysenck (1995)认为高创者由于过度包含的思维特点, 因此其注意过滤机制不像普通人那么严格, 这种过度包含性可能是一种抑制的失败, 是精神病患者、高精神质得分个体、创造性个体和天才具有的相似特征。高创造性个体有稳定的认知去抑制特点, 但是其与精神病患者不同, 在于高创造性个体能够拒绝不合适的反应, 而精神病患者则不能。如果是功能失调的认知去抑制类型,虽然也可能导致个体产生持续的想法, 但个体对已偏离最初观点的想法却无能为力, 而以抑制不相关反应的能力为基础的良好的认知抑制能力可能更有利于促进新颖的和独创的观点产生(Benedek et al., 2012)。因此, 这种认知抑制可能是一种功能良好的认知去抑制类型, 该类型支持产生流畅的、新颖的观点而不是产生僵硬的、病态持续的观点。而且, 有研究发现, 虽然精神分裂症与创造性思维得分呈正相关, 但是精神分裂症和创造性思维与认知抑制的关系不同(Rominger, Fink, Weiss, Bosch, & Papousek, 2017)。Rominger等人(2017)采用听觉抑制任务发现, 创造性思维与更高的认知抑制能力相关, 而精神分裂症与较低的认知抑制能力相关。高创者能够更好地抑制任务中不相关的听觉信息, 也可以解释为他们在创造性观点产生过程中更集中注意, 结果进一步支持抑制能力在创造性思维中起重要作用。
显然, 本研究结果与Dorfman等(2008)和Vartanian等(2007)的研究结果不一致。上述两个研究均采用了负启动任务作为有干扰的认知抑制任务, 结果发现, 创造性思维水平与不一致条件下的反应时呈正相关。抑制先前相关信息可能确实有利于创造性思维, 以更好地思考和整合对目前任务有用的大量信息。这与前人关于创造性思维与减少了的潜在抑制相关的观点一致(Carson et al., 2003; Eysenck, 1995)。导致与本研究结果不一致的原因可能是由于抑制概念的区别, 负启动任务和潜在抑制任务均是抑制前摄干扰或分心干扰而不是抑制优势反应的倾向(Friedman & Miyake, 2004)。
实验2成功操纵了时间压力情境, 并进一步采用皮肤电活动变化这一指标对认知抑制与创造性思维的关系进行检验。反应时结果表明, 高创者在有时间压力条件下的干扰效应量显著小于无时间压力条件下, 而低创者在有和无时间压力条件下的干扰效应量无显著差异, 表明时间压力在认知抑制与创造性思维的关系中起调节作用。面对时间压力时, 个体会想方设法在限定时间内完成任务, 所以有时间压力条件下, 个体的流畅性得到很大提高, 因而创造性也会得到提高, 所以相比无时间压力条件, 高创者在有时间压力条件下的干扰效应量更小, 表现出更高的认知抑制能力。这一结果还可以用注意力集中模型来解释(Karau & Kelly, 1992)。该模型认为, 时间压力会使个体的注意变窄, 与任务相关的特征凸显出来, 而与任务不十分相关的特征受到忽视。正如前面讨论中提到的, 高创者面对不同时间压力的任务情境, 能够调节自己的注意更好地适应所面对的情境, 使注意更集中于所解决任务的相关特征, 而更有效地抑制干扰的特征, 因此高创者比低创者的认知抑制更灵活。研究结果支持适应性认知抑制假说。
自主唤醒主要与腹内侧前额叶皮层、前扣带皮层、杏仁核和丘脑相关(Patterson, Ungerleider, & Bandettini, 2002), 同时也有研究发现背部前扣带皮层在抑制方面起着至关重要的作用(MacDonald, Cohen, Stenger, & Carter, 2000)。抑制需要心理努力并激发外周神经系统的交感活动增强。实验2的皮肤电结果发现, 高创者在颜色命名任务中不一致条件下的皮肤电活动变化显著高于一致条件, 而低创者在颜色命名任务中一致和不一致条件下的皮肤电活动变化皆没有显著差异。结果表明, 高创者在不同抑制条件下表现出生理唤醒水平的变化性, 而低创者则在各种条件下表现出基本一致的生理唤醒水平。这也符合Martindale (1999, 2007)的理论假设, 高创者会根据任务要求的不同而变换其生理唤醒水平, 支持创造性思维的适应性认知抑制假说。其他生理指标的研究也支持高创者比低创者具有更大的生理变化性。高创者完成仅需要创造性的任务中表现出大量α波活动, 而在仅需要智力的任务中表现最少的α波活动, 低创者在不同任务中表现出基本一致的α波活动水平(Martindale, 1999)。Kwiatkowski (2002)的研究也发现, 高创者和低创者在完成oddball任务中表现不同, 高创者在右半球的激活(P300幅度)比左半球更高, 而低创者在右半球的激活比左半球更低。
此外, 虽然在皮肤电活动变化指标上发现时间压力情境与创造性组别的交互作用达到边缘显著, 但是简单效应分析并没有发现高创者或低创者在有和无时间压力条件下的皮肤电活动变化差异显著, 但是低创者在有时间压力条件下的皮肤电活动变化比无时间压力条件下有升高的趋势(见图5)。这可能是因为相比无时间压力条件下, 低创者在有时间压力条件下更不能适应任务, 需要更大的抑制能力。间接的证据来自于α波同步化的研究。研究者倾向认为, α波同步化增加被认为是为了达到任务相关的内部定向注意而对外部刺激进行的自上而下的控制(Fink & Benedek, 2014)。谷传华等人(2015)的研究发现, 在表现高状态社会创造性时, 高创造特质的被试比低创造特质的被试出现更高的α波同步化, 同时还发现, 低创造特质被试在表现高状态社会创造性时, 右脑比左脑出现了更高的α波同步化。表明低创造特质被试在表现高状态创造性时, 需要更强大的抑制能力。研究者指出, SCR还可能反映无意识的评价过程, 也可能反映无意识的情感加工(Dawson et al., 2011)。实验2中在有时间压力条件下, 个体需要尽快做出反应, 否则会出现“反应太慢”的反馈。如果低创者在完成反应之后得到了不好的反馈, 那么其可能会对下一次反应预期一个消极结果, 或认为未来任务是一个不确定的情况, 所以可能导致其在有时间压力情境下比无时间压力情境下的皮肤电活动变化更高。本研究没有得到有显著差异的结果, 未来研究可以进一步加大样本量进行验证。
本研究的局限性是高创造性思维水平的个体选取的是在托兰斯创造性思维测验上得分高的被试, 而没有涉及其他领域。有研究发现, 认知抑制能力与艺术创造力之间呈现负相关(程丽芳, 胡卫平, 贾小娟, 2015)。这与本研究结果是不一致的。不一致的原因可能是因为认知抑制与创造性思维的关系在不同领域中的表现可能不尽相同。创造的心理加工非常复杂, 需要多个脑区协同工作。研究发现, 左额叶会抑制一般人(非艺术工作者)图形创造性思维任务中右半球的活动, 而且通过练习技巧消除这种抑制或通过对左额叶的特殊损伤可以促进艺术创造力(Huang et al., 2013)。也就是说左额叶的抑制作用使个体在图形创造力中表现更佳, 但如果某种原因导致左额叶的抑制作用降低则可以促进艺术创造力。由此可见, 不同的抑制程度可能确实对不同领域的创造力的影响不同。未来研究还需要进一步设计精巧的实验验证领域在认知抑制与创造性思维关系中所起的调节作用。
此外, 本研究也存在一些不足。在选取高低创造性思维水平被试时, 仅采用托兰斯创造性思维测验的词汇卷和图画卷, 测验较单一。未来研究可以采用多个测验的结果来进行综合评定。例如, 有研究采用《可能用途测验》和《托兰斯创造力测验》, 然后将这两个测验测量的独创性和流畅性维度分数转化后的Z分数之和作为衡量个体创造性思维水平的指标(Rominger et al., 2017)。其次, 本研究仅从发散思维角度对创造性思维水平进行考察, 而没有涉及到聚合思维。一般来说, 创造性思维包括发散思维和聚合思维两种基本形式。未来研究可以结合发散思维测验、聚合思维测验和智力测验等多个测验的项目来综合衡量个体的创造性思维水平, 以更全面地考察个体的创造能力(沃建中, 陈婉茹, 刘杨, 林崇德, 2010)。最后, 本研究仅将被试划分了高创组和低创组, 没有设置中等创造性思维水平组, 无法分析个体完成抑制任务时交感功能是哪一组的变化引起的, 抑或是两组共同作用之和。未来研究可以进一步增加中等创造性思维水平组, 为研究结果的可靠性提供更有利的证据。
5 结论
在本实验条件下, 可得出以下结论:(1)总体而言, 相比低创者, 高创者的认知抑制能力更高, 能够有效抑制优势的但不相关的反应倾向; (2)时间压力在认知抑制与创造性思维的关系中起调节作用, 高创者面对不同任务要求能够灵活调整自身的认知抑制水平, 其在有时间压力任务情境下的抑制能力高于无时间压力任务情境下, 且表现出变化的生理唤醒水平。结果支持创造性思维的适应性认知抑制假说。
参考文献
Creative and analytic thinkers differ in their use of attentional resources
Creativity under the gun
Model-based analysis of skin conductance responses: Towards causal models in psychophysiology
DOI:10.1111/j.1469-8986.2012.01483.x
URL
PMID:23094650
[本文引用: 1]

The empirical investigation of unobservable psychological processes usually rests on operational definitions. As an alternative, we propose the use of explicit causal models. This is particularly useful in psychophysiology, where formal models can be expressed mathematically, exploiting biophysical constraints, and inverted to yield estimates of unobservable processes. In psychophysiology, recent advances have been made in causal modeling for skin conductance responses, which we discuss to exemplify the development of such models. Empirical evidence suggests that these methods have a greater validity compared to operational approaches. This review concludes by considering the theoretical implications for the field of psychophysiology and benefits for practical data analysis.
The inhibitory mechanism of individuals with different scientific creativity
不同科学创造力个体干扰抑制机制的比较
The role of associative and executive processes in creative cognition
Differential effects of cognitive inhibition and intelligence on creativity
DOI:10.1016/j.paid.2012.04.014 URL [本文引用: 6]
To create or to recall? Neural mechanisms underlying the generation of creative new ideas
Intelligence, creativity, and cognitive control: The common and differential involvement of executive functions in intelligence and creativity
Publication recommendations for electrodermal measurements. Society for psychophysiological research Ad Hoc committee on electrodermal measures
DOI:10.1111/j.1469-8986.2012.01384.x
URL
PMID:7291438
[本文引用: 1]
This committee was appointed by the SPR Board to provide recommendations for publishing data on electrodermal activity (EDA). They are intended to be a stand-alone source for newcomers and experienced users. A short outline of principles for electrodermal measurement is given, and recommendations from an earlier report (Fowles et090009al., ) are incorporated. Three fundamental techniques of EDA recording are described: (1) endosomatic recording without the application of an external current, (2) exosomatic recording with direct current (the most widely applied methodology), and (3) exosomatic recording with alternating current090000to date infrequently used but a promising future methodology. In addition to EDA recording in laboratories, ambulatory recording has become an emerging technique. Specific problems that come with this recording of EDA in the field are discussed, as are those emerging from recording EDA within a magnetic field (e.g., fMRI). Recommendations for the details that should be mentioned in publications of EDA methods and results are provided.
Personality, Creativity and Latent inhibition
DOI:10.1002/per.572
URL
[本文引用: 1]
The current study set out to investigate the relationship between creativity, multi-dimensional schizotypy and personality more generally. This was achieved by analysing scores on a range of personality scales and measures of creativity, where it was found that the creativity measures were more closely related to asocial-schizotypy than positive-schizotypy. The study also sought to test Eysenck's prediction ([ 1993 ], [ 1995 ]) that, given the putative relationship between creativity and psychosis-proneness, high psychosis-prone scoring individuals and high creativity scoring individuals would demonstrate the same cognitive style of overinclusiveness on latent inhibition. However, the results failed to demonstrate any evidence of a shared widening of the associative horizon between high creativity and high psychosis-prone scorers. The findings are discussed in relation to multi-dimensional schizotypy. Copyright 2006 John Wiley & Sons, Ltd.
Decreased latent inhibition is associated with increased achievement in high-functioning individuals
Cognitive inhibition and artistic creativity: The moderating effect of cognitive style
认知抑制对艺术创造力的影响: 认知风格的调节作用
DOI:10.16187/j.cnki.issn1001-4918.2015.03.05
URL
[本文引用: 1]
以114名大学生为被试,采用Mittenecker指向测验和粘贴画任务考察了认知抑制能力与艺术创造力的关系,并采用镶嵌图形测验考察了认知风格在其中所起的调节作用。研究结果表明:(1)认知抑制能力与艺术创造力之间呈现负相关,认知抑制对于个体的创造程度、沟通传播水平和艺术创造综合印象可以起到显著的负向预测作用,对于个体艺术创造的可爱程度和想象水平可以起到边缘显著的负向预测作用;(2)认知风格在认知抑制能力与艺术创造力的关系中起着调节作用,主要表现为认知抑制能力对于场依存个体的创造程度、想象水平和沟通传播水平具有显著的预测作用,对于场独立个体的艺术创造力则不具有预测作用。
The different role of cognitive inhibition in early versus late creative problem finding
DOI:10.1037/aca0000036
URL
[本文引用: 2]
Abstract Previous research has suggested that ideas generated late in the creative process might require more executive control than those generated earlier. This in turn leads to the prediction that cognitive inhibition might play one role early in the process but a different role late in the process. The present investigation tested this prediction using a test of creative problem finding. Low cognitive inhibition was expected to facilitate an associative mode of processing, whereas high cognitive inhibition was expected to enable a deliberate, systematic mode of processing. An experiment involving 70 undergraduate students indicated that individuals' cognitive inhibition was correlated with fluency and flexibility, but not originality, on the problem-finding tasks. An interaction indicated that low cognitive inhibition enhanced originality initially, but later in the process, high cognitive inhibition was beneficial. Limitations of this investigation and future directions are explored.
Study of latent inhibition at high-level creative personality: The link between creativity and psychopathology
DOI:10.1016/j.sbspro.2012.01.142 URL [本文引用: 1]
The combined effects of neurostimulation and priming on creative thinking. A preliminary tDCS study on dorsolateral prefrontal cortex
Relationship between employee’s innovation (creativity) and time management
DOI:10.1016/j.sbspro.2011.10.541
URL
PMID:17054907
[本文引用: 2]
The article investigates the relationship between time management behaviours and attitudes with measures of creativity, as assessed by self-rated creativity and a measure of creative personality. Additionally, total creativity is examined, as the sum of the two creativity constructs when z-scored. Using data from a survey of 216 participants, results suggest that creativity is positively related to daily planning behaviour, confidence on long-range planning, perceived control of time and tenacity and negatively related to preference for disorganization. These results have theoretical implications for understanding how creativity relates to time management. Implications of the results are considered and future research directions identified.
The skin conductance response, anticipation, and decision-making
DOI:10.1037/a0022619 URL [本文引用: 2]
Up- and down regulating facial disgust: Affective, vagal, sympathetic, and respiratory consequences
DOI:10.1016/j.biopsycho.2005.02.006 URL [本文引用: 1]
Creativity and speed of information processing: A double dissociation involving elementary versus inhibitory cognitive tasks
DOI:10.1016/j.paid.2007.12.006 URL [本文引用: 3]
Creativity and the Stroop interference effect
DOI:10.1016/j.paid.2014.05.009
URL
[本文引用: 6]
Creative potential has been variably associated with disinhibition and defocused attention, focused attention and effective cognitive control, or a flexible adaption of cognitive control. The present study examined the relationship between creativity and cognitive control in a sample of design students and a control group. Cognitive control was assessed by a modified Stroop color naming task, in which two color words and two font colors were used to generate congruent and incongruent conditions. Design students showed stronger cognitive control as indicated by the absence of a Stroop interference effect, and performed generally better (faster) on the Stroop task than students in the control group did. Moreover, correlational analyses revealed associations between stronger cognitive control and higher scores in originality, fluency and ideational behavior in psychometric creativity tasks. These results suggest that one cognitive feature of creative individuals is effective suppression of dominant but irrelevant response tendencies.
EEG alpha power and creative ideation
The relations among inhibition and interference control functions: A latent-variable analysis
DOI:10.1037/0096-3445.133.1.101 URL [本文引用: 1]
Investigating the prosocial psychopath model of the creative personality: Evidence from traits and psychophysiology
DOI:10.1016/j.paid.2016.03.081 URL [本文引用: 1]
Effort intensity: Some insights from the cardiovascular system
In R. M. Ryan (Ed.),
Creativity and cognitive control: explorations of generation and evaluation skills
DOI:10.1207/S15326934CRJ152&3_09 URL [本文引用: 10]
Brain correlates underlying social creative thinking: EEG Alpha activity in trait vs. state creativity
社会创造性的脑机制: 状态与特质的EEG α波活动特点
The development of cognitive inhibition: Theories, definitions, and research evidence
In F. N. Dempster & C. J. Brainerd (Eds.),
DOI:10.1016/B978-012208930-5/50007-6
URL
[本文引用: 1]
This chapter illustrates the development of cognitive inhibition using theories, definitions, and research evidence. Several contemporary developmental theories emphasize the role of changes in inhibitory efficiency in accounting for developmental changes in other aspects of cognition and behavior. A vital assumption of such approaches is that inhibitory mechanisms become more proficient during early development and decline in efficiency in later development. This chapter examines the tenability of the hypothesis that certain forms of cognitive inhibition improve in efficiency over childhood. It begins with a discussion on cognitive inhibition in the context of historical and contemporary inhibition models. It examines some of the key characteristics of the cognitive inhibition construct. It is focused on distinguishing between behavioral and cognitive inhibition and between inhibition and interference. Finally, it also examines intentional and automatic forms of cognitive inhibition as well as a review of recent research evidence concerning developmental changes in these variables.
The influence of cognitive inhibition to creative scientific problem finding: Mediating effect of field cognitive style
认知抑制对创造性科学问题提出的影响:认知风格的中介作用
Evidence for a left-over-right inhibitory mechanism during figural creative thinking in healthy nonartists
DOI:10.1002/hbm.v34.10 URL [本文引用: 1]
The relationship between intelligence and creativity: New support for the threshold hypothesis by means of empirical breakpoint detection
DOI:10.1016/j.intell.2013.03.003 URL [本文引用: 1]
The road to creative achievement: A latent variable model of ability and personality predictors
DOI:10.1002/per.v28.1 URL [本文引用: 1]
The effects of time scarcity and time abundance on group performance quality and interaction process
DOI:10.1016/0022-1031(92)90045-L URL [本文引用: 1]
Autonomic arousal in cognitive conflict resolution
DOI:10.1016/j.autneu.2006.09.004
URL
PMID:17067858
[本文引用: 5]
Although cognitive efforts were reported to elicit global autonomic arousal, which cognitive processes associate with autonomic arousal has not been clear. We investigated autonomic arousal using event-related skin conductance responses (SCRs) during the Stroop color-word task. After baseline SCR deflections were determined in each trial block, SCRs were compared between cognitive conflict conditions (incongruent vs. congruent stimuli), between tasks assigned (word reading vs. color naming), and between erroneous and correct responses. Baseline SCRs were significantly greater at the beginning of each trial block. SCRs were significantly greater with incongruent than congruent stimuli while SCRs differed little between word reading and color naming. SCRs were greater when responses were incorrect. The results suggested that autonomic arousal occurs during cognitive conflict resolution in addition to mental set adoption for a task and in error awareness.
Individual differences in the neurophysiology of creativity (Unpublished doctorial dissertation)
Cognitive load and autonomic response patterns under negative priming demand in depersonalization-derealization disorder
DOI:10.1111/ejn.13183 URL
Dissociating the role of the dorsolateral prefrontal and anterior cingulate cortex in cognitive control
DOI:10.1126/science.288.5472.1835 URL [本文引用: 1]
Biological basis of creativity
In R. J. Sternberg (Ed.),
Creativity, primordial cognition, and personality
DOI:10.1016/j.paid.2007.05.014
URL
[本文引用: 2]
Potential creativity measured by the Alternate Uses Test, remoteness of word associations, and rated creativity of fantasy stories was found to be related to primordial content in the fantasy stories. Psychoticism and extraversion have been found to be related to creativity. There are theoretical reasons to think that they might also be related to use of primordial cognition. Potential creativity and primordial content were correlated with both extraversion and psychoticism. Potential creativity and primordial cognition were much more strongly related to extraversion than to psychoticism. The common trait binding these traits together may be disinhibition. Relationships among creativity, primordial cognition, and extraversion are much stronger among men than among women.
Enhancing verbal creativity: Modulating creativity by altering the balance between right and left inferior frontal gyrus with tDCS
Task-independent functional brain activity correlation with skin conductance changes: An fMRI study
DOI:10.1006/nimg.2002.1306 URL [本文引用: 1]
Behavioral inhibition and electrodermal activity during deception
DOI:10.1037//0022-3514.49.5.1427
URL
PMID:4078683
[本文引用: 1]
We tested the assumption that the act of inhibiting ongoing behavior requires physiological work. In a guilty knowledge test (GKT) paradigm, subjects were induced to attempt to deceive the experimenter on two separate occasions while electrodermal activity was measured. For 20 of the 30 subjects, overt behaviors (changes in eye movement and facial expression) were recorded during the second GKT. Results indicated that the incidence of behaviors decreased during their deceptive responses. This behavioral inhibition coincided with increases in skin conductance level. In addition to suggesting nonverbal correlates of deception, the results indicate that long-term behavioral inhibition may be a factor in psychosomatic disease.
Latent inhibition and openness to experience in a high-achieving student population
DOI:10.1016/S0191-8869(99)00101-4 URL [本文引用: 2]
Openness and extraversion are associated with reduced latent inhibition: Replication and commentary
DOI:10.1016/S0191-8869(02)00004-1 URL [本文引用: 3]
Power spectral density analysis of electrodermal activity for sympathetic function assessment
DOI:10.1007/s10439-016-1606-6 URL [本文引用: 1]
The role of (dis)inhibition in creativity: Decreased inhibition improves idea generation
DOI:10.1016/j.cognition.2014.09.001 URL [本文引用: 1]
Allusive thinking (remote associations) and auditory top-down inhibition skills differentially predict creativity and positive schizotypy
DOI:10.1080/13546805.2016.1278361 URL [本文引用: 2]
Creative motivation: Creative achievement predicts cardiac autonomic markers of effort during divergent thinking
DOI:10.1016/j.biopsycho.2014.07.010 URL [本文引用: 2]
Investigating in creativity
DOI:10.1037/0003-066X.51.7.677 URL [本文引用: 1]
Forgetting as a consequence and enabler of creative thinking
Studies of interference in serial verbal reactions
DOI:10.1037/h0054651 URL [本文引用: 1]
Toward a psychology of chronic time pressure: Conceptual and methodological review
Cognitive disinhibition and creativity
Variable attention facilitates creative problem solving
Creative potential, attention, and speed of information processing
DOI:10.1016/j.paid.2007.04.027
URL
[本文引用: 2]
Despite the conceptual overlap between intelligence and creativity, little systematic work exists on the link between creativity and speed of information processing. We hypothesized that differential focusing of attention determines the relationship between creative potential and speed of information processing. Supporting our predictions, we found negative correlations between creative potential and reaction time on tasks not involving interference (Hick Task, Concept Verification Task), but positive correlations between creative potential and reaction time on tasks requiring the inhibition of interfering information (Negative Priming, Global Precedence). This pattern of results suggests that the relation between creative potential and reaction time is a function of the differential focusing of attention.
The eye movements differences during category learning process between high and low creativity students
创造能力不同学生的分类加工过程差异的眼动特点
The relationship between creative thinking and cognitive inhibition
创造性思维与认知抑制的关系
Cross cultural comparison of creative thinking between Chinese and American students using Torrance Test
《托兰斯创造性思维测验》(TTCT)的测试和中美学生的跨文化比较
Creativity as flexible cognitive control
DOI:10.1037/a0017379
URL
[本文引用: 2]
Creative individuals have been described in terms suggestive of greater automatic processing (e.g., defocused attention, looser associations) and greater controlled processing (e.g., greater abilities to focus while working on a creative task). Both views cannot be correct from a static ability-related perspective. On the other hand, both views could be correct if creative individuals are better able to modulate the functioning of their cognitive control system in a context-sensitive manner. The present study (N 50) assessed individual differences in creativity in terms of original responses on the Torrance Test of Creative Thinking (Torrance, 1974) and also in terms of creative behavior on the Creative Achievement Questionnaire (Carson, Peterson, & Higgins, 2005). The same participants performed a color ord Stroop task. Creative individuals were neither more nor less capable of overriding cognitive conflicts on incongruent (relative to congruent) Stroop trials. On the other hand, creative individuals displayed more flexible cognitive control, as defined by greater cognitive control modulation from trial to trial. Implications for theories of creativity and its underlying processing basis are discussed.
Patterning and nonpatterning in creative cognition: Insights from performance in a random number generation task
DOI:10.1037/a0025452 URL [本文引用: 3]
Short-term attentional perseveration associated with real-life creative achievement
DOI:10.3389/fpsyg.2013.00191
URL
PMID:3635848
[本文引用: 3]
There are at least two competing hypotheses of how attention interacts with creative cognition, although they are not mutually exclusive. The first hypothesis is that highly creative people are particularly flexible at switching their attention – that is, they adaptively shift focus among different attentional levels using cognitive control. The second, less common, view is that creative people exhibit attentional persistence, or an ability for sustained attention. We suggest these two views need not be competing, as they may both operate, but on different time scales or on different components of creativity. In the present study we examined the role of attention in real-world creative achievement and in divergent thinking. In Experiment 1 participants with high and low real-world creative achievements identified whether the stimulus contained letters S or H within hierarchically constructed letters (e.g., large S made of small Es – global level; large E made up of small Ss – local level), which were presented in blocks of eight trials per level. In Experiment 2 participants with high, medium, and low creative achievements identified the same stimulus letters, but in blocks of five, seven, and nine trials per level. Results from both experiments indicated that people with high creative achievements made significantly more errors on trials in which they had to switch the level of attention, even after controlling for general intelligence. In Experiment 2, divergent thinking was also assessed, but it was not related to switching cost. Results from both experiments demonstrate that real-world creative acts relate to increased levels of attentional persistence, even if it comes with the cost of perseveration in certain circumstances.
The Differences between High Creative Persons and Low Creative Persons in Directing Forgetting Effects
高低创造性思维水平者定向遗忘效应的差异研究
Gray matter volume of the lingual gyrus mediates the relationship between inhibition function and divergent thinking
Standardization research on RAVEN’s standard progressive matrices in China
瑞文标准推理测验在我国的修订
The effects of three pressures and creative self-efficacy on creativity
三种压力与创意自我效能感对创造力的影响
The effects of emotional states on executive functioning
情绪唤起对执行功能的作用
Zooming into creativity: Individual differences in attentional global- local biases are linked to creative thinking
Effects of coping and cooperative instructions on guilty and informed innocents’ physiological responses to concealed information
DOI:10.1016/j.ijpsycho.2012.01.022 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}