

1 问题提出
青春期被概念化为第二个“敏感期”, 神经内分泌的发育变化增加其对环境的敏感性(Del Giudice et al., 2011), 面对压力性生活事件的增加, 尤其是涉及人际关系的事件, 青春期内外化问题发生率不断增加, 并对个性发展、学业成绩和身体健康产生负面影响(Madigan et al., 2016)。此外, 某些遗传特征也会影响青少年社会适应(见综述: Plomin et al., 2013)。社会适应(social adjustment)是一个广泛的概念, 涉及个体在社会环境中调整自己的行为、情感和认知, 以实现社会认可和期望的发展目标(Cavell, 1990)。社会适应涵盖适应能力和适应问题两个主要方面:适应能力包括积极主动、接纳合作、亲社会行为等; 适应问题包括外化问题, 如分心多动, 愤怒攻击等行为, 以及内化问题, 如抑郁、焦虑等(Chen & French, 2008)。尽管社会适应的发病风险受多种因素影响, 然而遗传与环境的相互作用对其的解释仍潜力十足, 并受到心理学、精神病学和表观遗传学研究者的重视(Hou et al., 2023; Mitchell et al., 2016)。鉴于此, 本研究通过多位点遗传变异的评估, 以及关键环境风险的测量, 旨在探讨遗传风险在不同环境压力形式(近端和远端压力)与青少年社会适应(抑郁症状、亲社会行为和自伤行为)之间的调节作用。
1.1 主要人际关系:亲子关系
Vrshek-Schallhorn等人(2015)提出在探究青少年社会适应的G × E效应时, 候选环境的选择是一个十分重要的环节。人际关系质量是影响个体社会适应能力的一个重要因素, 研究发现高遗传风险的青少年在主要人际压力源(而非次要人际或次要压力源)与社会适应之间表现出更强的关联(Starr & Huang, 2019)。亲子关系作为家庭生态环境中的核心元素, 一直是发展心理学、心理咨询与实践工作者研究的焦点(Bush & Peterson, 2013)。过往研究发现, 积极健康的亲子关系对个体的情感功能和适应能力产生显著的积极影响, 反之消极危险的亲子关系通常会导致心理健康问题(Lindell et al., 2021)。此外, 中国强调相互依存的文化价值观, 例如亲缘关系与人际和谐(Greenfield et al., 2006), 因此, 亲子关系对青少年社会适应的影响更大(Wang et al., 2021)。不仅如此, 在一项探究亲子关系与孩子社会适应的元分析中, Van Dijk等人(2020)发现亲密感、安全依恋和积极的亲子沟通等高品质的亲子关系可以提高孩子自尊、生活满意感; 而家长控制、亲子冲突则显著预测孩子内外化问题行为。
1.2 环境与环境的交互作用
需要澄清的是, 压力敏感性模型(stress sensitization model)提出, 早期压力暴露可能会加剧个体对未来压力刺激的敏感性, 从而影响青春期社会适应, 这种现象可被形容为“环境与环境” (environment × environment, E × E)之间的相互作用。相关研究表明童年逆境(如儿童期受虐、父母丧失、父母分居)可能增加个体对后续近端压力的敏感性, 增加诸如抑郁(Stroud, 2019)、失眠(Turner et al., 2020)、自伤行为(Campos et al., 2020)和攻击性行为(Caruso et al., 2021)等社会适应风险。在一项调查童年期逆境与成人精神障碍的大样本研究(N = 34653)中, 结果显示相比没有遭受过童年逆境的个体, 具有童年逆境史的人群近端压力性事件与抑郁、焦虑、创伤后应激障碍的相关更大(McLaughlin et al., 2010)。不仅如此, 经历过儿童期虐待的青少年在面对近期较低强度压力时也有着更高的抑郁风险(La Rocque et al., 2014)。与此同时, Liu等人(2018)在其纳入71项研究数十万参与者的有关童年期创伤和自伤行为的系统综述与元分析中, 结果显示各个类型的童年期创伤均与自伤行为相关, 且这种关联在非临床样本中更加强烈。研究也发现遭受过童年期虐待和忽视的个体, 其照顾他人的能力也会降低, 变得不那么无私, 从而影响利他、道德和亲社会行为(Music, 2011)。
1.3 压力敏感性的遗传变异
大量研究证实了遗传因素在青少年社会适应发生发展中的影响, 其中遗传变异增加暴露于童年期逆境或近端压力源后患抑郁症的风险(Assary et al., 2018), 并且会影响亲社会行为(Music, 2011; Si et al., 2020)。然而, 考虑到不同发育阶段应激压力的交互影响, 简单的遗传与环境(gene × environment, G × E)模型难以满足青少年社会适应的复杂病因。因此, 研究者提出了一个更复杂的模型, 即遗传变异可以调节童年期逆境所产生的压力敏感性, 青少年社会适应的差异是“遗传−环境−环境gene × environment × environment” G × E × E三者交互作用的结果, 其中第一个“E”指童年期逆境, 第二个“E”则指近端压力源(Keers & Pluess, 2017)。在一项调查抑郁的G × E × E效应的研究中, 结果显示血清素转运蛋白基因, 儿童期受虐和成人创伤经历的三者交互作用可以显著预测抑郁的产生(Grabe et al., 2012)。有多种生物学机制可用以解释遗传风险如何影响由童年期逆境导致的个体压力敏感性, 举例来说, 儿童期逆境可能导致压力相关系统和结构的发育发生变化, 例如下丘脑−垂体−肾上腺(hypothalamic-pituitary-adrenal, HPA)轴的敏感性增加和海马体积减少(Cicchetti & Rogosch, 2012), 而基因则可能对这些系统的可塑性变化产生影响, 青少年在经历了儿童期逆境后更容易出现与压力相关的神经回路的持久性改变(Heim et al., 2008)。
1.4 超越单一候选基因:多位点方法
差别易感性模型(the differential susceptibility, Belsky et al., 2007)提出“可塑性”基因会产生近似于“近朱者赤, 近墨者黑”的效应, 即具有某种基因的个体在不利环境下可能增加风险, 但在优势环境中则可能预测积极结果。因此, 对于在较低童年逆境中成长的青少年, 遗传的“风险”可能预示着积极的应对策略和其他弹性因素, 从而有助于缓解儿童期逆境后出现抑郁症的风险。这些观点也得到现有研究的支持(Schuler et al., 2017; Vinkers et al., 2015), 研究指出遗传变异可以通过G × E × E交互效应来调节早期逆境对近端压力敏感性的影响, 并主要集中在HPA轴和血清素(serotonin)系统。在一项探究与HPA轴系统相关的促肾上腺皮质激素受体基因(corticotrophin hormone receptor, CRHR1)调节由童年期逆境导致的压力敏感性的纵向研究中, 结果显示CRHR1位点A等位基因与更大的压力敏感性相关(Starr et al., 2014)。然而, 相比于单基因受到的较大争议, 多位点遗传谱评分(multilocus genetic profile scores, MGPS)通过提取重要遗传位点中的未加权风险等位基因的累积得分(Starr & Huang, 2019), 具有更大检验功效以及更稳健特性, 还可以避免跨多个基因的多次测试, 从而减少I型错误的可能性(Zhang & Belsky, 2020), 对于涉及多个基因和环境交互的复杂性状, MGPS考虑了多个位点的遗传信息, 更能够应对复杂遗传结构。在一项对超过7000名参与者出生队列样本中, Keers和Pluess (2017)发现较高的MGPS可以调节童年期经济条件(住房、社会阶层等)导致的成年后个体压力敏感性。
1.5 HPA轴遗传变异
HPA轴系统是主要的应激反应生理系统之一, HPA轴系统功能的失调与紊乱可以显著影响个体情绪, 记忆以及睡眠等生物反应, 因此一直被视为抑郁、自伤和亲社会行为的重要候选基因(Keers & Pluess, 2017; Starr & Huang, 2019)。研究发现童年逆境可以影响HPA轴生理功能上的变化如皮质醇水平、应激反应性以及昼夜节律等(Gunnar & Quevedo, 2007; Stroud et al., 2016), 而且这些变化在经历近期应激暴露之后, 会导致抑郁风险上升(Schuler et al., 2017)。在一项探究HPA轴系统FKBP5基因累积遗传风险与创伤暴露对抑郁影响的前瞻性研究中, Zimmermann等人(2011)发现FKBP5基因MGPS与创伤暴露可以显著预测青少年抑郁症, 而且这一发现在荷兰样本中得到了重复(Vinkers et al., 2015)。与此同时, 有研究发现盐皮质激素受体(肾上腺皮质产生的一类激素)基因CA单倍体个体, 在低父母养育压力水平时表现出更低的亲社会行为(Endedijk et al., 2019)。不仅如此, 在一项纳入7项研究(包括3585名参与者)探究应激发生的遗传标志物新近系统综述中, 结果显示HPA轴, 血清素和催产素系统多基因累加遗传风险可以预测个体社会心理易感性, 并调节近端生活事件所产生的影响(Bahji et al., 2021)。此外, 研究也证实HPA轴MGPS能够调节早期逆境和近期压力对社会适应的影响, 从而影响诸如自伤(Hou et al., 2023)、亲社会行为(Donovan & Henley, 2010)。
1.6 目前的研究
不同于以往遗传与环境的G × E研究范式, 我们探究新兴的G × E × E三者交互效应对青少年社会适应的影响(见图1)。其次, 过往大多数的G × E研究都严重依赖素质−压力模型(diathesis-stress model)的观点, 因此, 往往选择消极环境因素(儿童期虐待、父母离异或负性生活事件)探究其对消极心理因素如抑郁、焦虑、愤怒或攻击行为等的影响。相比之下, 当前的研究通过引入不同类型环境因素(童年期创伤和亲子关系)拟利用差别易感性框架来检验HPA轴系统多基因敏感性是否会调节不同环境因素对青少年社会适应的影响。换句话说, 本研究假设基因更敏感的青少年会由于更差的环境(童年期创伤高分和亲子关系低分)产生更多的抑郁症状和自伤行为, 也会因为更积极的环境(高水平的亲子关系和低水平低童年期创伤)表现出更多的亲社会行为。通览HPA轴系统基因和社会适应的相关研究, 本研究使用MGPS来表示一组候选基因中变异的累加效应。最后, 之前大多数涉及青少年社会适应的G × E研究都是横向的, 本研究使用追踪数据以降低共同方法偏差和有偏回忆问题。
图1
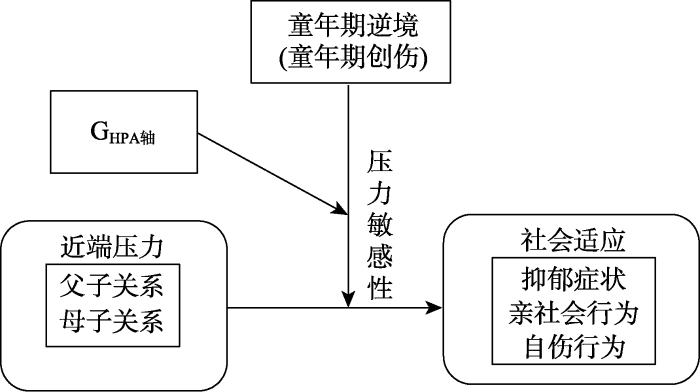
本研究包含主实验和补充实验两个独立但相互关联的组成部分, 以更全面地探究青少年社会适应与遗传、环境之间的关系。主实验(遗传与环境交互作用传统研究范式:问卷调查)旨在通过量表测量青少年社会适应的多个方面, 包括抑郁症状、亲社会行为和自伤行为, 这些指标不仅是遗传与环境研究中经典且常见的测量因素, 还在HPA轴系统的调控下发挥关键作用, 同时也是青少年心理健康的综合反映(指标选择原因见网络版附录1)。通过问卷调查, 能够获取受试者的主观报告, 从而深入了解其社会适应水平及其在不同方面的表现。补充实验采用行为学任务, 以客观、量化的方式测量青少年在抑郁症状和亲社会行为方面的表现。旨在弥补问卷调查可能存在的主观因素, 并提供更客观、直接的观察指标, 以增强对青少年社会适应的全面理解。
总之, 本研究利用纵向数据, 拟检验青少年多基因风险得分作为压力敏感性与青少年社会适应之间的调节因素。预计随着遗传风险评分的增加, 童年期逆境和近端压力之间的交互作用将会增加, 且与差别易感性假说一致, 多基因敏感性水平高且具有高水平亲子关系的青少年有着更低的抑郁症状、自伤行为以及更高的亲社会行为, 反之亦然。
2 方法
2.1 研究程序和对象
研究对象来源于湖南某中学初二学生, 按班级采取整群抽样, 所有项目均获得学校、学生及其家长的知情同意, 首次测量为2021年11月第三周的工作日下午进行, 测量近端压力因素(亲子关系), 由统一培训的心理学教师和研究生担任主试, 前往调研学校, 并保证每个班级两名主试配合, 班主任在旁协助调查, 共收集899份问卷数据。此外, 本次也实施基因采集(口腔上皮细胞), 在收集前委托班主任告知学生, 在采集前30 min每名学生使用清水漱口, 之后不能进食或喝饮料等影响样本纯度行为。采集过程中按班级(50人左右)分配严格训练的主试人员, 不少于7人, 采集过程约30 min, 共获得785份基因样本(由于有些同学拒绝参与)。采集完成后的口腔刮棒立即采用冰袋寄回某生物科技公司, 随后进行DNA提取与纯化, 并对全部DNA进行琼脂糖凝胶电泳质检, 结果显示均在10ng/ul左右, 满足iMLDR多重SNP分型技术要求。全样本均无缺失基因型, 后使用阴性对照检查分型结果, 基因分型效率良好(DNA提取和分型过程见网络版附录2)。需要澄清的是为使得基因采集顺利, 本次测量过程问卷测试在问卷星平台发布在学校机房进行, 相关研究表明传统纸质测量与线上问卷不存在较大差异, 符合测量不变性要求(陈永泰, 何有世, 2008; 于洪彦, 黄晓治, 2011), 基因采集在机房隔壁由生物公司工程师指导布置的短期实验室进行。
追踪为2022年10月第三周的工作日下午, 测量远端压力因素(儿童期虐待)和青少年社会适应(抑郁症状, 亲社会行为和自伤行为), 测量过程同前, 本次测试采用纸质问卷测量。后经过匹配具有追踪和基因数据的参与者共有700名(首次测量参与者年龄14.15 ± 0.63岁), 其中女生320 (45.7%)名, 男生380 (54.3%)名; 独生子女323 (46.2%)名, 非独生子女377 (53.8%); 父母婚姻状况:初婚583 (83.3%)名, 离异28 (4.0%)名, 再婚48 (6.8%)名, 其他类型41 (5.9%)名; 家庭经济水平:很好125 (17.9%)名, 较好414 (59.0%)名, 一般148 (20.2%)名, 较差8 (1.2%)名, 很差5 (0.7%)名。追踪样本与实际纳入样本人口学特征均不存在显著差异(χ2s < 9.91, ps > 0.053), 两次调查不存在结构化流失。由于第二次使用纸质问卷, 因此存在一定缺失值, 通过缺失值分析结果显示:变量缺失0, 不完整个案75 (10.71%), 值缺失238 (0.65%), 缺失率较小, 判定为随机缺失。本研究使用的缺失数据插补方法为:EM插补(expectation maximization imputation)。EM假设数据缺失类型为随机缺失, 并使用当前数据信息和指定模型对缺失数据进行“最佳猜测” (Bennett, 2001), 适用于连续变量。
2.2 研究工具
2.2.1 亲子关系
使用Buchanan等人(1991)编制亲子亲密度量表(Parent-child Intimacy Questionnaire), 分别对父子和母子的关系进行评述, 各9个条目(如, 当你向你父/母亲表露情感时, 你感觉很舒服、自然?), 采用5点评分, 从1为“完全不符合”到5为“完全符合”。量表得分越高表示青少年与父亲(或母亲)的关系越紧密, 可以适用中国青少年群体(胡义秋 等, 2023)。本研究中父子关系部分的Cronbach’s α系数为0.92, 母子关系部分Cronbach’s α系数为0.92。量表结构效度良好, 父子部分验证性因素分析的拟合指数:χ2 = 60.96, df = 27, CFI = 0.96, TLI = 0.95, RMSEA = 0.06, SRMR = 0.03; 母子部分验证性因素分析的拟合指数:χ2 = 100.74, df = 27, CFI = 0.92, TLI = 0.89, RMSEA = 0.07, SRMR = 0.05。配对样本t检验得分显示母子关系得分显著高于父子关系(M ± SD父子 = 30.51 ± 9.36, M ± SD母子 = 33.18 ± 8.59, t = 10.49, p < 0.001)。
2.2.2 童年期创伤
使用Aslund等人(2011)自编的儿童期虐待量表(Childhood abuse scale)用以评估青少年童年期创伤经历, 在获得其本人的正式批准回复后, 由两名英语专业博士采用互译法翻译成中文, 并接受由心理学教授、博士以及一线心理学教师组成的10人专家团进行评估, 在删除两项不符合中国语境条目后, 最终量表共包含7个题项(如:您的父母是否曾推搡或殴打您, 或对您使用任何其他形式的暴力?从不或很少(0), 至少每年一次(1)), 总分范围为0~7分。本研究儿童期受虐的Cronbach’s α系数为0.72, 验证性因素分析结果显示:χ2 = 26.53, df = 14, CFI = 0.90, TLI = 0.95, RMSEA = 0.08, SRMR = 0.06。
2.2.3 抑郁症状
采用Andresen等人(1994)编制的简版流调中心抑郁量表(Short Form of the Center for Epidemiological Studies Depression Scale, SF-CES-D), 在我国青少年群体中有较好的适用性(Yang et al., 2018), 为单维量表共包含10个题目(如, 我感到忧郁), 采用4级(0~3)评分, 该量表的总分在0~30分之间。本研究中简版流调中心抑郁量表的Cronbach’s α系数为0.85, 验证性因素分析的拟合指数:χ2 = 103.07, df = 35, CFI = 0.90, TLI = 0.89, RMSEA = 0.08, SRMR = 0.07, 量表结构效度良好。
2.2.4 亲社会行为
采用由寇彧等人(2007)修订的Carlo和Randall (2002)编制的亲社会倾向量表(Prosocial Tendencies Measures, PTM)中文版用以测量青少年亲社会行为水平。量表包含6个维度, 共包含26个项目(如, 有人在场时, 我会竭尽全力帮助别人), 采用5点记分, 从1表“非常不像我”到5表“非常像我”, 在我国青少年群体有着较好的适用性(Xie et al., 2016)。本研究中量表Cronbach’s α系数为0.96, 验证性因素分析的拟合指数:χ2 = 546.89, df = 299, CFI = 0.91, TLI = 0.90, RMSEA = 0.04, SRMR = 0.04, 量表有着较好的结构效度。
2.2.5 自伤行为
采用简版自我伤害问卷(Deliberate Self-Harm Inventory, DSHI; Gratz, 2001)用以测量参与者的自我伤害行为, 该量表在中国青少年中表现出良好的信效度(胡义秋 等, 2023), 为单维量表, 有9个条目(如, 用烟头、打火机或火柴烫自己), 从0表示“没有”到5表示“5次及以上”, 得分越高表明自伤行为越严重。调查完成后课题组随机挑选10名参与者对问卷进行评价, 均未出现严重不适感。本研究中简版自我伤害问卷的Cronbach’s α系数为0.85。量表结构效度良好, 验证性因素分析的拟合指数:χ2 = 142.34, df = 27, CFI = 0.94, TLI = 0.92, RMSEA = 0.08, SRMR = 0.04。
2.3 分析方法
数据可用性分析:问卷共同方法偏差检验以及对HPA轴系统多基因进行Hardy-Weinberg平衡检验考察基因取样情况。
多位点遗传谱评分(multilocus genetic profile scores, MGPS)计算与相关分析:基于之前的研究和生物环境敏感性理论(theory of biological sensitivity; Ellis & Boyce, 2008), 筛选被认为可能与青少年社会适应相关的遗传基因, 最终选定HPA轴系统相关基因SKA2、NR3C1、NR3C2、FKBP5作为候选基因。之后在国际人类基因组计划提供的单核苷酸多态性(single nucleotide polymorphisms, SNP)公共数据库(
表1 HPA轴系统相关SNP与其功能的关系以及编码方案 (具体描述见网络版附录3)
| SNP | 与功能的关系 | 风险等位基因 |
|---|---|---|
| SKA2 rs7208505 | Valenzuela-García等人(2023)发现自杀受害者中rs7208505等位基因A的比例高于健康对照组。 | A |
| NR3C1 rs6198 | Rovaris等人(2013)发现C等位基因可以显著预测每天吸烟数量和尼古丁依赖测试总分。 | C |
| NR3C1 rs41423247 | 根据Pagliaccio等人(2014)研究, G等位是影响HPA轴的风险等位基因。 | G |
| NR3C2 rs17581262 | 有研究发现rs17581262 的G等位基因与较小的海马和杏仁核体积, 以及与遭受过儿童期受虐的个体中较高的重症抑郁风险相关(Gerritsen et al., 2017)。 | G |
| FKBP5 rs1360780 | 根据Pagliaccio等人(2014)研究, T等位基因是影响HPA轴的风险等位基因。 | T |
| FKBP5 rs9470080 | 相比健康对照组, 功能性癫痫发作和重度抑郁症患者的T等位基因显著增加(Asadi- Pooya et al., 2023)。 | T |
相关分析与G × E × E效应检验:使用相关分析探究各核心变量之间的相关; 探究青少年社会适应的G × E × E效应; 最后简单斜率图以及Johnson-Neyman检验(Hayes & Matthes, 2009)探究MGPS具体调节效应。
遗传与环境交互作用理论模型验证:为厘清上述基因和环境交互作用所属的理论模型, 采用显著性区域(region of significance, RoS, Roisman et al., 2012)分析法分别考察HPA轴系统MGPS与环境(亲子关系和儿童期虐待)对青少年社会适应的影响, 以判断是否符合素质−压力模型或是差别易感性模型。相较传统的检验方法, 显著性区域分析法具有诸多优势:首先, 通过将环境变量(X)的最佳显著区间(regions of significance on X, RoS on X)确定在M ± 2 SD以内(覆盖正态分布样本95%的数据)。其次, 采用量化标准对模型进行判断:1)交互作用比例指标(the proportion of interaction index, PoI)表示的良好交互效应(b)占全部交互效应(b+w)的百分比。通常, 若PoI趋于0, 则表示完全符合素质−压力模型, 当0.40 ≤ PoI ≤ 0.60说明符合差别易感性模型(王美萍, 2015); 2)受影响比例指标(the proportion affected index, PA)表示受到良好交互效应影响的参与者百分比。通常PA的取值范围在0~0.5间, 若PA值近似于0, 则说明交互作用模式符合素质−压力模型, 而大于0.16靠近0.50 (0.40~0.60)则符合差别易感性模型。此外, 还需要考虑交叉点(cross-over point)所处位置, 即两条回归线交叉点所对应的横坐标位置。最后, 通过检验X2 (或ZX2)对Y的预测是否显著, 用以确定变量间是否存在非线性关系, 以排除遗传与环境的交互作用误判为差别易感性模型的可能。
3 结果
3.1 数据可用性检验
本研究数据均来自被试自我报告, 无法规避存在的共同方法偏差问题。使用Harman单因子法对6个问卷的所有题项进行探索性因素分析, 采用最优斜交法的主成分分析法。分析结果显示, 共有13个因子的特征根大于1, 第一个因子的方差解释率为15.81% (低于临界指标40%)。此外, 打包各个量表3个因子载荷最大的条目作为对应的潜变量指标, 使用Mplus进行单因子模型的验证性因素分析, 结果显示模型拟合很差:χ2 = 695.30, df = 135, CFI = 0.49, TLI = 0.42, RMSEA = 0.18, SRMR = 0.16。上述结果提示不存在严重的共同方法偏差问题。
表2 SNP数据
| 基因 | 位点 | 等位基因 | MAF | MAF (1000g- CHBS) | 基因型和分布 | Hardy-Weinberg equilibrium | |||
|---|---|---|---|---|---|---|---|---|---|
| 11 | 12 | 22 | χ2 | p | |||||
| SKS2 | rs7208505 | G > A | 0.40 | 0.40 | 252 (36.0%) | 341 (48.7%) | 107 (15.3%) | 0.23 | 0.63 |
| NR3C1 | rs6198 | T > C | 0.0002 | 0.001 | 698 (99.7%) | 2 (0.03%) | 0 (0%) | 0.001 | 0.97 |
| rs41423247 | G > C | 0.22 | 0.22 | 431 (61.6%) | 236 (33.7%) | 33 (4.7%) | 0.01 | 0.92 | |
| NR3C2 | rs17581262 | A > G | 0.19 | 0.19 | 464 (66.3%) | 210 (30.0%) | 26 (3.7%) | 0.14 | 0.71 |
| FKBP5 | rs1360780 | C > T | 0.30 | 0.29 | 344 (49.1%) | 287 (41.0%) | 69 (9.9%) | 0.64 | 0.42 |
| rs9470080 | C > T | 0.35 | 0.34 | 292 (41.7%) | 325 (46.4%) | 83 (11.9%) | 0.26 | 0.61 | |
注:11 = 主要等位基因构成的纯合子频率, 12 = 杂合子频率, 22 = 次要等位基因构成的纯合子频率。
对所有基因位点进行Hardy-Weinberg遗传平衡检验(见表2), 基因型观测值与期望值较为吻合, 结果符合Hardy-Weinberg平衡定律。不仅如此, 本次研究中次要等位基因频率(Minor Allele Frequency, MAF)大部分大于0.05 (除rs6198), 而且与千人基因组计划中中国北京汉族人群最小等位基因频率MAF (1000g-CHBS)类似, 结果显示本次取样较好。
3.2 MGPS评分计算与相关分析
表3 描述统计量及相关分析结果
| 变量 | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| 父子关系 | 1 | ||||||
| 母子关系 | 0.72** | 1 | |||||
| 童年期创伤 | −0.26** | −0.20** | 1 | ||||
| 抑郁症状 | −0.45** | −0.39** | 0.27** | 1 | |||
| 亲社会行为 | 0.14** | 0.15** | −0.15** | −0.09* | 1 | ||
| 自伤行为 | −0.19** | −0.14** | 0.36** | 0.34** | −0.17** | 1 | |
| MGPS | 0.04 | 0.08* | −0.03 | 0.03 | 0.01 | 0.01 | 1 |
| M ± SD | 30.51 ± 9.36 | 33.18 ± 8.59 | 0.93 ± 1.40 | 17.70 ± 6.01 | 96.42 ± 17.63 | 2.56 ± 6.26 | 4.04 ± 1.67 |
注:1为父子关系; 2为母子关系, 3为童年期创伤; 4为抑郁症状; 5为亲社会行为; 6为自伤行为; 7为HPA轴系统MGPS。
*p < 0.05, **p < 0.01, ***p < 0.001, 下同。
3.3 青少年社会适应的G × E × E检验
接下来, 考察HPA轴系统遗传变异在青少年童年期创伤经历后的压力敏感性对社会适应影响的调节作用。基于三因素之间的交互作用, 使用SPSS 27.0宏程序PROCESS v3.4 Model 3进行检验, 所有连续变量标准化后, 将所有主效应以及二者和三者之间的交互作用同时输入模型, 结果如表4和5所示。仅母子关系不能有效预测青少年自伤行为, 而亲子关系和童年期创伤在三个方程中均可以显著预测青少年抑郁(|βs亲子关系| > 0.10, ps < 0.01; |βs童年期创伤| > 0.14, ps < 0.001)。此外, 在抑郁症状方程中, 三者交互作用显著(β父子 = 0.09, p = 0.003; β母子= 0.07, p = 0.03), 同时HPA轴系统MGPS和母子关系与童年期创伤的交互作用也显著(βsMGPS > 0.07, ps < 0.048; β = 0.09, p = 0.01)。在亲社会行为方程中, 母子关系模型三者交互作用显著(β = −0.07, p = 0.047), 亲子关系和童年期创伤的交互作用显著(β父子 = −0.09, p = 0.01; β母子 = −0.08, p = 0.03)。虽然自伤行为方程中未发现G × E × E交互作用, 但童年期创伤和MGPS的交互作用也可以(边缘)预测青少年自伤行为(β父子 = 0.09, p = 0.02; β母子 = 0.07, p = 0.05), G × E × E 效应约占青少年社会适应方差的1% (ΔR2抑郁症状 = 0.01, p = 0.001; ΔR2亲社会行为 = 0.01, p = 0.05; ΔR2自伤行为 = 0.003, p = 0.95)。
表4 青少年社会适应的G × E × E检验父子模型
| 变量 | 抑郁症状 | 亲社会行为 | 自伤行为 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β | t | p | 95% CI | β | t | p | 95% CI | β | t | p | 95% CI | |
| 父子关系 | −0.40 | −11.66*** | < 0.001 | (−0.47, −0.34) | 0.11 | 2.77** | 0.01 | (0.03, 0.18) | −0.10 | −2.82** | 0.005 | (−0.18, −0.03) |
| 童年期创伤 | 0.18 | 4.88*** | < 0.001 | (0.11, 0.25) | −0.15 | −3.78*** | < 0.001 | (−0.23, −0.07) | 0.32 | 8.31*** | < 0.001 | (0.24, 0.40) |
| MGPS | 0.07 | 1.98* | 0.048 | (0.001, 0.13) | −0.01 | −0.14 | 0.89 | (−0.08, 0.07) | 0.03 | 0.71 | 0.48 | (−0.05, 0.10) |
| E1 × E2 | 0.03 | 0.85 | 0.39 | (−0.04, 0.09) | −0.09 | −2.55* | 0.01 | (−0.16, −0.02) | −0.04 | −1.20 | 0.23 | (−0.11, 0.03) |
| E1 × G | −0.01 | −0.29 | 0.78 | (−0.07, 0.05) | 0.02 | 0.54 | 0.59 | (−0.05, 0.09) | 0.001 | 0.03 | 0.98 | (−0.07, 0.07) |
| E2 × G | 0.05 | 1.50 | 0.13 | (−0.02, 0.12) | −0.001 | −0.03 | 0.98 | (−0.08, 0.08) | 0.09 | 2.39* | 0.02 | (0.02, 0.16) |
| E1 × E2 × G | 0.09 | 3.00** | 0.003 | (0.03, 0.15) | −0.04 | −1.29 | 0.20 | (−0.11, 0.02) | 0.01 | 0.19 | 0.85 | (−0.06, 0.07) |
| R2 | 0.24 | 0.04 | 0.15 | |||||||||
| F | 31.06*** | 4.56*** | 17.27*** | |||||||||
注:E1为亲子关系; E2为童年期创伤; G为MGPS, 加粗字体表示显著, 下同。
表5 青少年社会适应的G × E × E检验母子模型
| 变量 | 抑郁症状 | 亲社会行为 | 自伤行为 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β | t | p | 95% CI | β | t | p | 95% CI | β | t | p | 95% CI | |
| 母子关系 | −0.36 | −10.31*** | < 0.001 | (−0.43, −0.29) | 0.13 | 3.39*** | < 0.001 | (0.05, 0.20) | −0.07 | −1.89 | 0.06 | (−0.14, 0.003) |
| 童年期创伤 | 0.23 | 6.32*** | < 0.001 | (0.16, 0.30) | −0.14 | −3.71*** | < 0.001 | (−0.22, −0.07) | 0.34 | 9.16*** | < 0.001 | (0.27, 0.41) |
| MGPS | 0.08 | 2.17* | 0.03 | (0.02, 0.15) | −0.02 | −0.55 | 0.58 | (−0.10, 0.05) | 0.02 | 0.55 | 0.58 | (−0.05, 0.09) |
| E1 × E2 | 0.09 | 2.62** | 0.01 | (0.01, 0.14) | −0.08 | −2.21* | 0.03 | (−0.15, −0.01) | −0.03 | −0.86 | 0.39 | (−0.10, 0.04) |
| E1 × G | −0.03 | −0.86 | 0.39 | (−0.09, 0.04) | −0.04 | −1.16 | 0.25 | (−0.11, 0.03) | −0.02 | −0.68 | 0.50 | (−0.09, 0.04) |
| E2 × G | 0.02 | 0.62 | 0.53 | (−0.05, 0.09) | −0.02 | −0.53 | 0.60 | (−0.10, 0.06) | 0.07 | 1.93 | 0.05 | (−0.001, 0.15) |
| E1 × E2 × G | 0.07 | 2.16* | 0.03 | (0.01, 0.13) | −0.07 | −1.99* | 0.047 | (−0.13, −0.001) | −0.02 | −0.47 | 0.64 | (−0.08, 0.05) |
| R2 | 0.21 | 0.05 | 0.14 | |||||||||
| F | 26.10*** | 5.22*** | 16.47*** | |||||||||
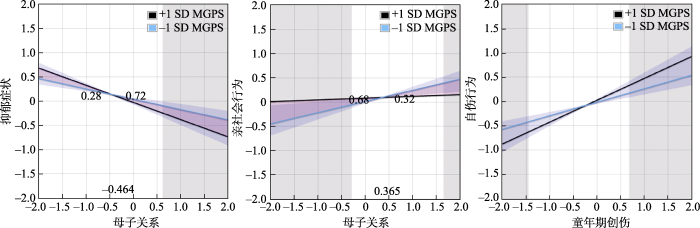
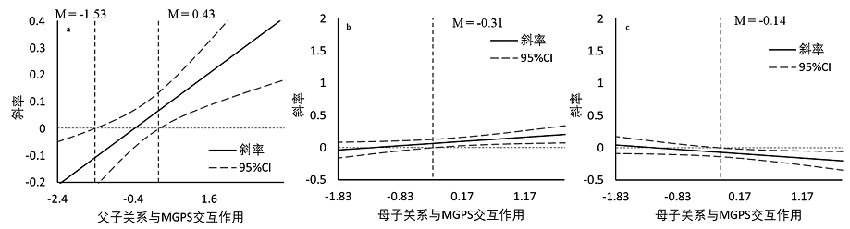
简单斜率图结果:图2a显示对于那些低HPA轴系统MGPS的青少年, 父子关系与童年期创伤时的抑郁症状均显著相关(βLCA = −0.35, p < 0.001; βHCA = −0.46, p < 0.001); 且对于那些高HPA轴系统MGPS青少年来说, 父子关系与低童年期创伤时的抑郁相关增加(β = −0.49, p < 0.001), 但当遭受高童年期创伤, 父子关系与抑郁症状相关性降低(β = −0.30, p < 0.001)。且Johnson-Neyman (见网络版附录4图a)检验结果显示, 当HPA轴系统MGPS标准差小于−1.53时, 父子关系与MGPS交互作用为负数且持续下降, 当HPA轴系统MGPS标准差大于−1.53但小于0.43时, 父子关系和MGPS交互作用不显著, 而当HPA轴系统MGPS标准差大于0.43时, 父子关系和MGPS交互作用为正数且持续增加。作为这种效应的另一种解释, 我们检查了近期压力暴露的遗传调节是否因远端压力暴露而强化(即相同G × E × E交互作用的另一种分解, E1为童年期创伤, E2为亲子关系)。结果显示, G × E交互作用在不同童年期创伤水平下差异显著(β低童年期创伤 = −0.07, p = 0.05; β高童年期创伤 = 0.08, p = 0.04), 结果进一步支持遗传因素在不同压力条件下的作用差异。
图2

图2
青少年社会适应G × E × E的简单斜率图
注:a为青少年抑郁症状方程父子模型, b为抑郁症状方程母子模型, c为亲社会行为方程母子模型以及d为Johnson-Neyman 斜率图; L-LCA for low MGPS - low childhood abuse即低HPA轴MGPS和低童年期创伤, L-HCA for low MGPS -high childhood abuse即低HPA轴MGPS和高童年期创伤, H-LCA for high MGPS - low childhood abuse即高HPA轴MGPS和低童年期创伤, H-HCA for high MGPS -high childhood abuse即高HPA轴MGPS和高童年期创伤。
图2b显示对于那些低HPA轴系统MGPS青少年来说, 母子关系与童年期创伤时的抑郁症状均显著相关(βLCA = −0.34, p < 0.001; βHCA = −0.31, p < 0.001); 且对于那些高HPA轴系统MGPS青少年来说, 母子关系与低童年期创伤时的抑郁症状相关性增加(β = −0.49, p < 0.001), 但当遭受高童年期创伤, 母子关系与抑郁症状相关性降低(β = −0.23, p < 0.001)。且Johnson-Neyman (见网络版附录4图b)检验结果显示当HPA轴系统MGPS标准差大于−0.31时, 母子关系与MGPS交互作用显著且持续增加。作为这种效应的另一种解释, 我们检查了近期压力暴露的遗传调节是否因远端压力暴露而强化。结果显示, G × E交互作用在不同童年期创伤水平下差异显著(β低童年期创伤 = −0.08, p = 0.04; β高童年期创伤 = 0.10, p = 0.03)。
此外, 图2c显示对于那些低HPA轴系统MGPS青少年来说, 母子关系与童年期创伤时的亲社会行为相关, 而在高低童年期创伤水平之间无显著差异(βLCA = 0.17, p = 0.01; βHCA = 0.15, p = 0.01); 但对于那些高HPA轴系统MGPS青少年来说, 母子关系可以显著预测低童年期创伤时的亲社会行为(β = 0.19, p = 0.003), 但与遭受高童年期创伤时的亲社会行为无关(β = −0.06, p = 0.40)。Johnson- Neyman检验结果(见网络版附录4图c)显示, 仅当HPA轴系统MGPS标准差大于−0.14时, 母子关系与MGPS的交互作用开始显著。作为这种效应的另一种解释, 我们也检查了近期压力暴露的遗传调节是否因远端压力暴露而强化。结果显示, G × E交互作用仅在高童年期创伤水平下边缘显著(β 低童年期创伤 = 0.003, p = 0.94; β高童年期创伤 = −0.11, p = 0.02)。
尽管在青少年自伤行为方程中并未发现G × E × E三者交互作用, 但在父子关系模型中也发现了HPA轴系统MGPS和童年期创伤的二元交互作用。因此, 使用PROCESS Model 1仅检验HPA轴系统MGPS在童年期创伤对青少年自伤行为影响中的单调节作用, 结果显示童年期创伤与HPA轴系统MGPS的交互作用显著(β = 0.09, p = 0.02, 具体回归系数表见网络版附录5), 并且G × E (童年期创伤)效应约占青少年自伤行为方差的1% (ΔR2 = 0.01, p = 0.02)。随后的Johnson-Neyman 检验(见图2d)结果显示当HPA轴系统MGPS标准差大于−2.29时, 童年期创伤可以预测青少年自伤行为, 具体表现为随着MGPS得分的增加, 童年期创伤对青少年自伤行为的负向预测增加, 斜率增大。
根据Dick等人(2015)的建议, 作为稳健性测试, 检验包括性别、是否独生子女、父母婚姻状况以及家庭经济地位作为协变量的后续模型。在这些模型中, 当满足以下条件时, (a)包含这些协变量的主要影响, 只有性别(β抑郁 = 1.07, p < 0.001)和父母婚姻状况(βs > 0.47, ps < 0.02)可以影响青少年社会适应和(b)对于双向G×协变量、亲子关系×协变量和童年期创伤×协变量交互作用(共计12个附加协变量, 仅有抑郁症状方程中亲子关系和性别的交互作用显著β = −0.03, p < 0.001), 三者(二者)交互作用仍然显著(β抑郁症状父子 = 0.10, p = 0.001; β 抑郁症状母子 = 0.08, p = 0.01; β亲社会行为 = −0.08, p = 0.02; β 自伤行为 = 0.09, p = 0.001)。这表明即使考虑了这些协变量, 遗传和环境交互作用对青少年社会适应仍然具有显著影响。
3.4 遗传与环境交互作用理论模型验证
仅考察已经发现的遗传与环境的交互作用(亲子关系对抑郁、母子关系对亲社会行为以及童年期创伤对自伤行为)。结果见表6所示, 母子关系与MGPS的交互作用可以显著预测青少年抑郁症状和亲社会行为(β抑郁症状 = −0.07, p = 0.04; β亲社会行为 = −0.10, p = 0.01), 此外童年期创伤和MGPS的交互作用也可以预测青少年自伤行为(β = 0.09, p = 0.01)。其次, 采用RoS方法进一步考察HPA轴系统MGPS与环境的交互模式(见表7和图3), 结果显示三个方程中, 不论环境类型(母子关系/童年期创伤), 还是心理结果变量类型(抑郁症状、亲社会行为和自伤行为), PoI指数在0.32~0.72之间, 且PA也在0.36~0.68之间, 近似为0.5。此外, X2和ZX2在预测青少年社会适应不显著, 可以排除非线性关系。综上所述, HPA轴系统MGPS与环境的交互模式支持差别易感性模型。
表6 遗传与环境交互作用
| 结果 | 抑郁症状 | 亲社会行为 | 自伤行为 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 预测 | 父子关系 | 母子关系 | 母子关系 | 儿童期虐待 | ||||||||||||
| β | t | p | 95% CI | β | t | p | 95% CI | β | t | p | 95% CI | β | t | p | 95% CI | |
| 环境 | −0.35 | −9.76*** | < 0.001 | (−0.42, −0.28) | −0.28 | −7.77*** | < 0.001 | (−0.35, −0.21) | 0.13 | 2.47*** | 0.01 | (0.03, 0.24) | 0.36 | 10.28*** | < 0.001 | (0.29, 0.43) |
| MGPS | −0.04 | −1.08 | 0.28 | (−0.11, 0.03) | −0.03 | −0.90 | 0.37 | (−0.10, 0.04) | 0.04 | 0.69 | 0.49 | (−0.07, 0.14) | 0.02 | 0.57 | 0.57 | (−0.05, 0.09) |
| G × E | −0.01 | −0.27 | 0.79 | (−0.07, 0.06) | −0.07 | −2.08* | 0.04 | (−0.14, −0.004) | −0.10 | −2.17* | 0.03 | (−0.18, −0.01) | 0.09 | 2.49* | 0.01 | (0.02, 0.15) |
| R2 | 0.12 | 0.09 | 0.03 | 0.14 | ||||||||||||
| F | 32.78*** | 22.83*** | 5.77*** | 36.62*** | ||||||||||||
表7 遗传与环境交互作用ROS检验结果
| 结果变量 | 参数 | 环境变量的显著区间 | POI | PA | X2/ZX2 | 交叉点 | |
|---|---|---|---|---|---|---|---|
| 下边界 | 上边界 | ||||||
| 抑郁症状 | 母子关系 × MGPS | −4.16 | 0.62 | 0.72 | 0.68 | ns. | −0.46 |
| 亲社会行为 | 母子关系× MGPS | −0.28 | 1.66 | 0.32 | 0.36 | ns. | 0.37 |
| 自伤行为 | 童年期创伤× MGPS | −1.44 | 0.68 | 0.62 | 0.59 | ns. | −0.23 |
图3
3.5 敏感性检验
3.5.1 单个基因位点与环境的交互作用对青少年社会适应的影响
探究每单个基因位点G × E × E对青少年社会适应影响。结果显示(具体结果见网络版附录6), 在青少年抑郁症状方程中, 仅有FKBP5基因rs1360780 (β = 0.10, p < 0.001)和rs9470080多态性(β = 0.14, p = 0.003)在父子关系模型中与环境的三者交互作用显著, 其他所有位点的遗传与环境交互作用不显著(|β|s < 0.10, ps > 0.09)。而在亲社会行为模型中, 不论是在父子关系还是母子关系模型中, 其他所有位点的遗传与环境交互作用不显著(|β|s < 0.10, ps > 0.06)。此外, 以相同的方式探究单个基因位点在儿童受虐对青少年自伤行为的影响中, 结果显示仅有FKBP5基因rs1360780多态性(β = 0.14., p = 0.01)与童年期创伤的交互作用显著, 其他所有位点的遗传与环境交互作用不显著(|β|s < 0.07, ps > 0.19)。此外, 网络版附录7探究了HPA轴单基因对青少年社会适应的主效应, 结果显示仅有NR3C1基因rs6198多态性对亲社会行为的主效应显著(β = 0.09, p = 0.02)。结果提示, 多基因累加得分相比单基因研究在抑郁遗传解释率方面更有优势。
3.5.2 独立的HPA轴系统(n-1)MGPSs与环境的交互作用对青少年社会适应的影响
采用单次拿出一个基因位点, 构建6个独立的HPA轴系统(n-1)MGPSs, 探究其与环境的交互对青少年社会适应的影响。结果显示在抑郁症状方程中, 仅在母子模型中拿出NR3C2基因rs17581262多态性后(β = 0.03, p = 0.12)三者交互作用不显著; 其他所有的独立(n-1)MGPSs与环境的三者交互作用均显著(|β|s > 0.04, ps < 0.04)。而在亲社会行为方程中, 仅在拿出SKA2基因rs7208505多态性后(β = −0.04, p = 0.12)三者交互作用不显著, 其他独立的MGPSs与环境的三者交互作用均(边缘)显著(|β|s > 0.04, ps < 0.05)。此外, 用相同的方式, 结果显示在自伤行为模型中, 6个独立(n-1)MGPSs与童年期创伤的交互作用均显著(|β|s > 0.04, ps < 0.047)。结果提示多基因累加得分相比单基因研究更加稳定, 具体结果见网络版附录8。
3.5.3 内部一致性分析
将总样本随机分为两个子样本, 子样本在全部人口学上没有差异(χ2s < 2.98, ps > 0.40,)。结果显示, 虽然样本量降低, 但G × E × E三者交互作用仍可以(边缘)预测青少年抑郁症状(β父子sample 1 = −0.01, p = 0.90; β父子sample 2 = 0.09, p = 0.07, β母子sample 1 = −0.01, p = 0.90; β母子sample 2 = 0.09, p = 0.07)和亲社会行为(βsample 1 = −0.08, p = 0.06; βsample 2 = −0.06, p = 0.32)。此外, 在青少年自伤行为模型中也发现了HPA轴系统MGPS与童年期创伤交互作用显著(βsample 1 = 0.06, p = 0.16; βsample 2 = 0.13, p = 0.01)。上述结果提示, 多基因遗传与环境交互作用对青少年社会适应的影响具有较好的稳定性和可靠性, 具体结果见网络版附录9。
4 补充研究:G × E × E效应对青少年社会适应影响的行为学研究证据
为进一步深入了解G × E × E效应对青少年社会适应的影响, 进行了一项补充实验。在这一节中, 将详细介绍实验的方法、参与者的特征, 以及两个关键行为学任务的设计和实施:情绪Stroop任务(抑郁症状)和独裁者游戏(亲社会行为)。
4.1 方法
4.1.1 参与者
研究对象来源于研究1参与者, 行为学实验于2023年6月进行, 通过实验传单招募研究1参与者135名, 所有参与者视力或矫正视力均正常, 无色盲或色弱, 实验结果后给予小礼物进行奖励。后经过匹配具有问卷、基因和实验数据的参与者共有126名, 其中女生62 (49.6%)名, 男生64 (50.4%)名; 独生子女68 (53.8%)名, 非独生子女58 (46.2%); 父母婚姻状况:初婚102 (80.7%)名, 离异8 (6.7%)名, 再婚10 (7.6%)名, 其他类型6 (5.0%)名; 家庭经济水平:很好27 (21.0%)名, 较好67 (52.9%)名, 一般31 (25.2%)名, 较差1 (0.8%)名, 很差0 (0%)名。研究1样本与补充研究纳入样本人口学特征均不存在显著差异(χ2s < 3.00, ps > 0.12)。以效应量为0.02, 本研究126名被试, 以α = 0.05为标准, 检验相关多位点遗传谱评分的交互效应, 其功效估计范围为50%。
4.1.2 材料与程序
由于仅在青少年抑郁症状和亲社会行为发现G × E × E效应, 因此仅设计抑郁症状(情绪Stroop任务)和亲社会行为(独裁者游戏)实验。具体如下:
情绪Stroop任务(emotional Stroop task):情绪Stroop范式中, 刺激呈现为带颜色的情绪词(积极、消极、抑郁)和中性词代替原始Stroop任务中的色词干扰(Myers & McKenna, 1996)。情绪Stroop效应表现为:被试对情绪词颜色的反应和中性词颜色的反应差异显著, 体现了情绪信息加工的影响。实验用E-prime 2.0软件编程, 在计算机上操作实施, 屏幕背景为灰色。具体程序如下(程序示意图见网络版附录10说明1):首先呈现注视点“+”300 ms, 随后呈现颜色词汇直到被试做出反应或2000 ms消失进入到下一试次。被试任务是对呈现词汇的颜色进行相应的按键反应:红色按“F”键、绿色按“J”键、蓝色按“K”键。实验共包括120个试次, 红、绿和蓝色词汇分别包括40个试次, 其中, 每种颜色的积极、消极、中性和抑郁词(情绪词从“汉语情绪词系统”中选取)各10个。正式实验前, 被试先进行15个试次的练习。记录参与者完成Stroop任务的反应时(参与者完成任务所需的时间, 即识别词语颜色的反应速度)和正确率(参与者在完成任务时正确识别词语颜色的比例)。
独裁者游戏(dictator games):参照国内外研究, 本研究采用独裁者游戏范式来测量个体的亲社会行为水平(陈思静 等, 2015; Raihani & Mcauliffe, 2014)。独裁者游戏由E-Prime 2.0程序编制及呈现, 屏幕背景为灰色。具体程序如下(程序示意图见网络版附录10说明2):首先在屏幕上呈现任务要求, 告知参与者“下面将进行一个在线分配代币的游戏, 由您和在另一个实验室的参加者共同分得一笔金钱(10个代币)。游戏中有两种角色, A分配者和B接受者。分配者有权以任何方式分配这笔钱, 接受者只能接受, 无权拒绝。在游戏中, 电脑程序会在您与另一个参加者之间随机分配角色。你们互不认识, 以后也不会相见, 分配结果将以匿名的方式呈现给对方, 并且最终的实验报酬将以分配的金额挂钩。获得的金额越大, 最终报酬就越多。”阅读完毕之后, 计算机呈现:“现在系统开始指定角色A和B, A有权提出分配, B无法拒绝该分配方案。”为了更加真实, 计算机屏幕会呈现挑选界面, 并告知被试为分配者, 并说明:“恭喜您成为角色A, 您可以根据自己的喜好来分配10个代币, 分配所获的收益将与最终实验报酬挂钩。”最后, 进入正式分配环节, 告知被试:“请在下面方框内输入给对方分配的金额数目。”以测量参与者亲社会行为。
为了营造真实的在线博弈情境, 实验对接受者设置了基本信息, 包括性别, 年龄, 职业, 共创造了八个接受者(即进行8次分配), 如第一个接受者编号253, 年龄24 岁, 职业是在校大学生。实验将对八个接受者信息在被试间实施平衡化处理。
4.1.3 分析方法
由于样本量的问题, 将亲子关系、童年期创伤和HPA轴系统MGPS以50%百分位数为分割点分为高低分组, 采用多因素方差分析探究G × E × E效应对青少年抑郁症状和亲社会行为的影响。
4.2 结果:G × E × E效应对青少年社会适应的影响
为检验G × E × E效应, 进行2 (HPA轴系统MGPS: 高低分组) × 2 (亲子关系: 高低分组) × 2 (童年期创伤: 高低分组)的多因素方差分析。表8结果显示对于所有情绪词的正确率, 母子关系主效应显著(Fs (1, 118) > 6.69, ps < 0.01, η2ps > 0.054), 母子关系高分组准确率显著高于低分组, 并未发现其他主效应或交互效应; 在反应时维度, 表9结果显示父子关系主效应显著(Fs (1, 118) > 6.69, ps < 0.01, η2ps > 0.034), 父子关系低分组在积极词汇和抑郁词汇的反应时显著高于高分组, 即低分组更容易受到情绪词的影响。此外, 在母亲模型中, G × E × E效应在总反应、积极词汇和消极词汇上显著, Fs (1, 118) > 3.64, ps < 0.04, η2p > 0.030。事后分析结果显示在高童年期创伤、高HPA轴系统MGPS条件下, 母子关系低分组在积极词汇和抑郁词汇的反应时显著高于高分组, 未发现其他主效应或交互效应。
表8 G × E × E检验对情绪Stroop任务正确率的影响
| 预测因素 | 总正确率 | 积极词 | 消极词 | 抑郁词 | 中性词 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | η2p | F | p | η2p | F | p | η2p | F | p | η2p | F | p | η2p | |
| 父子关系 | 2.16 | 0.14 | 0.018 | 2.63 | 0.11 | 0.022 | 2.38 | 0.13 | 0.020 | 2.04 | 0.16 | 0.017 | 1.45 | 0.23 | 0.012 |
| 童年期创伤 | 0.08 | 0.78 | 0.001 | 0.25 | 0.62 | 0.002 | 0.00 | 0.99 | 0.000 | 0.09 | 0.77 | 0.001 | 0.09 | 0.76 | 0.001 |
| MGPS | 2.41 | 0.12 | 0.020 | 2.88 | 0.09 | 0.024 | 2.99 | 0.08 | 0.025 | 1.82 | 0.18 | 0.015 | 1.82 | 0.18 | 0.015 |
| E1 ×E2 | 0.22 | 0.64 | 0.002 | 0.23 | 0.64 | 0.002 | 0.07 | 0.79 | 0.001 | 0.12 | 0.73 | 0.001 | 0.55 | 0.46 | 0.005 |
| E1 × G | 0.02 | 0.89 | 0.000 | 0.01 | 0.93 | 0.000 | 0.09 | 0.76 | 0.001 | 0.00 | 0.99 | 0.000 | 0.20 | 0.89 | 0.000 |
| E2 × G | 0.31 | 0.58 | 0.003 | 0.24 | 0.63 | 0.002 | 0.09 | 0.77 | 0.001 | 1.00 | 0.32 | 0.008 | 0.15 | 0.70 | 0.001 |
| E1 × E2 × G | 0.64 | 0.43 | 0.005 | 0.46 | 0.50 | 0.004 | 0.45 | 0.50 | 0.004 | 0.33 | 0.57 | 0.003 | 1.50 | 0.22 | 0.013 |
| 母子关系 | 7.75** | 0.01 | 0.062 | 7.71** | 0.01 | 0.061 | 6.69** | 0.01 | 0.054 | 8.06** | 0.01 | 0.064 | 7.56** | 0.01 | 0.060 |
| 童年期创伤 | 0.01 | 0.93 | 0.000 | 0.02 | 0.88 | 0.000 | 0.12 | 0.73 | 0.001 | 0.01 | 0.91 | 0.001 | 0.001 | 0.98 | 0.000 |
| MGPS | 3.02 | 0.09 | 0.025 | 3.50 | 0.06 | 0.029 | 3.66 | 0.05 | 0.030 | 2.51 | 0.12 | 0.021 | 2.20 | 0.14 | 0.018 |
| E1 × E2 | 0.87 | 0.35 | 0.007 | 1.04 | 0.01 | 0.009 | 0.47 | 0.49 | 0.004 | 0.69 | 0.41 | 0.006 | 1.28 | 0.26 | 0.011 |
| E1 × G | 0.83 | 0.36 | 0.007 | 0.72 | 0.40 | 0.006 | 0.78 | 0.35 | 0.007 | 0.84 | 0.36 | 0.007 | 0.80 | 0.37 | 0.007 |
| E2 × G | 0.21 | 0.64 | 0.002 | 0.16 | 0.69 | 0.001 | 0.04 | 0.85 | 0.000 | 0.76 | 0.38 | 0.006 | 0.12 | 0.73 | 0.001 |
| E1 × E2 × G | 0.43 | 0.51 | 0.004 | 0.41 | 0.53 | 0.003 | 0.30 | 0.59 | 0.002 | 0.13 | 0.72 | 0.001 | 1.11 | 0.30 | 0.009 |
表9 G × E × E检验对情绪Stroop任务反应时的影响
| 预测因素 | 总反应时间 | 积极词 | 消极词 | 抑郁词 | 中性词 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | η2p | F | p | η2p | F | p | η2p | F | p | η2p | F | p | η2p | |
| 父子关系 | 2.98 | 0.09 | 0.025 | 4.10* | 0.045 | 0.034 | 0.93 | 0.34 | 0.008 | 5.61* | 0.02 | 0.045 | 1.10 | 0.29 | 0.009 |
| 童年期创伤 | 1.45 | 0.23 | 0.012 | 0.31 | 0.58 | 0.003 | 0.94 | 0.33 | 0.008 | 2.33 | 0.12 | 0.019 | 1.76 | 0.19 | 0.015 |
| MGPS | 0.20 | 0.65 | 0.002 | 0.04 | 0.85 | 0.000 | 0.03 | 0.88 | 0.000 | 0.15 | 0.70 | 0.001 | 0.75 | 0.39 | 0.006 |
| E1 × E2 | 0.31 | 0.58 | 0.003 | 1.30 | 0.26 | 0.011 | 0.04 | 0.84 | 0.000 | 0.20 | 0.66 | 0.002 | 0.10 | 0.75 | 0.001 |
| E1 × G | 0.61 | 0.44 | 0.005 | 0.11 | 0.74 | 0.001 | 0.26 | 0.61 | 0.002 | 0.15 | 0.70 | 0.001 | 2.40 | 0.12 | 0.020 |
| E2 × G | 0.08 | 0.78 | 0.001 | 0.48 | 0.49 | 0.004 | 0.01 | 0.95 | 0.000 | 0.18 | 0.68 | 0.001 | 0.01 | 0.93 | 0.000 |
| E1 × E2 × G | 0.88 | 0.35 | 0.007 | 0.38 | 0.54 | 0.003 | 1.69 | 0.20 | 0.014 | 0.99 | 0.32 | 0.008 | 0.32 | 0.78 | 0.003 |
| 母子关系 | 1.62 | 0.21 | 0.014 | 1.34 | 0.25 | 0.011 | 0.29 | 0.59 | 0.002 | 3.32 | 0.07 | 0.027 | 1.24 | 0.27 | 0.010 |
| 童年期创伤 | 1.28 | 0.26 | 0.011 | 0.16 | 0.69 | 0.001 | 0.68 | 0.41 | 0.006 | 2.37 | 0.13 | 0.020 | 1.72 | 0.19 | 0.014 |
| MGPS | 0.39 | 0.54 | 0.003 | 0.23 | 0.64 | 0.002 | 0.17 | 0.68 | 0.001 | 0.17 | 0.68 | 0.001 | 0.90 | 0.34 | 0.008 |
| E1 × E2 | 0.25 | 0.62 | 0.002 | 0.24 | 0.62 | 0.002 | 0.01 | 0.91 | 0.000 | 0.28 | 0.60 | 0.002 | 0.72 | 0.40 | 0.006 |
| E1 × G | 0.52 | 0.48 | 0.004 | 0.69 | 0.41 | 0.006 | 0.42 | 0.52 | 0.004 | 0.45 | 0.50 | 0.004 | 0.28 | 0.60 | 0.002 |
| E2 × G | 0.09 | 0.77 | 0.001 | 0.40 | 0.53 | 0.003 | 0.003 | 0.96 | 0.000 | 0.14 | 0.71 | 0.001 | 0.002 | 0.97 | 0.000 |
| E1 × E2 × G | 3.64* | 0.05 | 0.030 | 4.61* | 0.03 | 0.038 | 4.56* | 0.04 | 0.037 | 1.90 | 0.17 | 0.016 | 2.03 | 0.16 | 0.017 |
然而在亲社会行为方面, 结果显示不论是父子关系还是母子关系模型均未发现显著主效应或交互效应, Fs (1, 118) < 1.72, ps > 0.19。具体结果见网络版附录11。
5 讨论
基于压力敏感性理论以及新兴G × E × E模型, 使用统计上更稳健的MGPS方法, 本研究通过引入关键遗传因素(HPA轴系统), 童年期逆境(童年期创伤)以及近端重要人际因素(亲子关系), 结果显示遗传因素可以调节早期逆境所带来的压力敏感性, 青少年社会适应是G × E × E三者交互作用的结果, 这些发现可能是对现行G × E研究报告不一致结果的一种潜在解释, 对于丰富和理解G × E在精神病理学发展和预防心理健康问题中的作用具有重要意义。
越来越多的研究表明, 早期不良的生活经历会导致儿童生理变化, 并且这种变化可能持续存在并增加终生不良健康结果的风险(Rnic et al., 2018; Schönfelder et al., 2019)。与过往研究一致, 本研究发现童年期创伤可以显著预测青少年社会适应。不仅如此, 动物和人类的神经生物学研究也发现童年期创伤与细胞老化、HPA轴以及相关神经递质系统的变化有关(Szyf, 2012), 甚至可以影响脑结构, 特别是海马体结构和功能的改变, 而且这种影响是终身持续(Bremner, 2006)。其次, 本研究结果也显示青少年阶段重要的人际关系类型——亲子关系——均可以显著预测青少年社会适应。过往研究结果显示, 高水平的亲子关系可以提升青少年学业表现、自尊和归属感, 也会降低相关心理问题和障碍的发生风险(Carmona-Halty et al., 2022)。
候选基因和环境交互作用对社会适应的影响一直受到对研究结果不可靠的争议, 促使人们在样本量, 关键候选基因以及重要环境的选择上一直争论不休(Hou et al., 2023; Starr & Huang, 2019)。使用多位点遗传谱评分方法, 本研究首次检验了HPA轴系统多基因在童年期创伤和亲子关系交互作用对青少年社会适应的调节作用。结果显示不论是简单斜率图还是Johnson-Neyman检验均显示, 随着HPA轴系统MGPS增加, 亲子关系与童年期创伤的交互作用增强, 即高遗传敏感性个体, 压力敏感性更强。我们的结果与Keers和Pluess (2017)先前报道的结果一致, 发现童年虐待与遗传敏感性评分之间存在显著的相互作用。具体来说, 对于那些遗传敏感性得分较低的人来说, 是否遭受儿童期虐待对近端压力对抑郁作用几乎没有影响。而遗传敏感性得分高且遭受过高水平童年期创伤的个体复原力功能最低(亲子关系对青少年抑郁显著预测降低, 且抑郁症状得分高), 而遗传敏感性得分高但未遭受儿童期虐待的青少年, 亲子关系可以显著预测社会适应。
其次, 虽然仅在母子关系模型中发现G × E × E三者交互作用对青少年亲社会行为的影响, 结果显示当HPA轴系统MGPS标准差大于−0.14时, 童年期创伤与母子关系的交互作用开始显著。具体效应与抑郁症状方程类似, 即低HPA轴系统多基因时, 母子关系与童年期创伤时的亲社会行为相关, 且高低童年期创伤水平无显著差异; 但对于那些高HPA轴系统MGPS青少年来说, 母子关系可以正向预测低童年期创伤时的亲社会行为, 但与遭受高童年期创伤时的亲社会行为无关, 即高遗传敏感性和高童年期逆境抵消了母子关系对青少年亲社会行为的正向影响。
母亲和父亲在与孩子相处的时间、活动类型以及互动风格上存在明显差异(Ferreira et al., 2016)。在亲子互动中, 研究表明母亲更倾向于进行口头教育, 而父亲则更多地展现出身体刺激和交流的特点(Carlo et al., 2011)。实际上, 有研究证实父母在影响孩子亲社会行为方面存在显著的差异(Lindsey et al., 2010)。不仅如此, 在中国传统家庭中父亲往往扮演严肃的“红脸”形象, 而母亲则更多是温和的“白脸”形象, 这可能也是造成在遗传与环境交互作用对青少年社会适应存在父母差异的原因(王美萍 等, 2019; 周雅 等, 2017)。此外, 虽然自伤行为方程中未发现G × E × E三者交互作用, 但随后的童年期创伤与HPA轴系统MGPS二元交互作用检验结果显示, 随着MGPS得分增加, 童年期创伤对青少年自伤行为的预测增加, 即消极环境增加了青少年自伤行为。
值得注意的是, 之前的许多G × E研究仅关注消极环境或逆境, 或者假设较少的消极环境就是积极环境, 如父母没有敌意就相当于父母温暖, 又或是认为低消极心理结果就是高水平发展, 如低抑郁症状就是积极发展(见综述: Duncan & Keller, 2011), 本研究考察亲子关系、童年期创伤与遗传的交互对社会适应(抑郁症状、亲社会行为和自伤行为)的影响。一方面简单斜率图得分显示高HPA轴得分、高亲子关系得分和低童年期创伤抑郁症状得分最低, 亲社会行为得分最高; 而高HPA轴得分、低亲子关系和高童年期创伤抑郁症状得分最高, 亲社会行为得分最低, 即高遗传敏感性个体受环境影响性最大。另一方面, RoS方法检验结果也显示三种社会适应结果均支持差别易感性模型(Belsky & Pluess, 2013)。
过去的文献尚未详细说明青少年遗传敏感性如何调节亲子关系与青少年社会适应之间的关联机制。然而, 通过对HPA轴神经生理学功能的全面审视, 可以提出一些合理的假设。HPA轴系统与奖励惩罚和感觉寻求相关(Matthys et al., 2013)。正如Belsky和Pluess (2013)指出, 在边缘系统, 尤其是在杏仁核中, HPA轴系统的表达和基因变异与社会适应有关。因此, 对于奖励(如父母的温暖和支持)以及惩罚(如父母的敌意和愤怒胁迫), 相较于其他青少年, 高遗传敏感性的青少年可能更容易受到环境的影响。此外, 另一个潜在机制可能涉及DNA甲基化以及某些环境刺激导致的基因表达变化。比如, Beach等人(2010)发现, 遭受过儿童期虐待的个体, 与情绪相关的DNA甲基化水平存在显著且持久的差异。因此, 需要进一步进行神经心理学和表观遗传学研究, 以更全面地了解遗传特征与压力敏感性在环境刺激对青少年心理适应影响的机制。
有研究人员注意到, 如果模型中未考虑相关协变量 × 基因和环境相互作用, G × E的效应可能是由于混杂因素驱动的, 而不是由特定的遗传与环境变量本身导致(见方法学综述: Keller, 2014)。为此在稳健性测试中, 我们测试了青少年性别、是否独生子女、父母婚姻状况和家庭经济地位与亲子关系、童年期创伤和多基因敏感性之间所有可能的双向交互作用。在所有包含协变量相互作用的分析中, G × E × E三者交互作用仍显著, 且仅有性别和父母婚姻状况具有统计显著性。除了考虑协变量的作用外, 行为遗传学家还指出在解释G × E排除基因与环境相关性的重要性(Dunn et al., 2011)。相关分析结果表明, HPA轴系统MGPS与仅与母子关系存在较小相关。不仅如此, 三种敏感性分析结果也显示多基因遗传研究具有更好的有效性和稳定性(Starr et al., 2021; Starr & Huang, 2019)。
为进一步验证G × E × E效应对青少年社会适应的影响, 我们通过两个经典且简单的行为实验作为补充证据。结果显示在情绪词的准确率和反应时上发现了亲子关系主效应, 且与过往研究类似高水平的亲子关系可以预测情绪处理与执行功能(Kim & Kochanska, 2012; Sabag-Cohen, 2009), 此外也在母子关系模型中发现了G × E × E三者交互作用。但仍需要指出的是, 在探究青少年亲社会行为的独裁者游戏中并未发现任何主效应或交互作用。其可能的原因是本研究中的独裁者游戏主要用于测查亲社会行为中的合作行为且测量过于简单。事实上, 传统的亲社会研究可能无法探测青少年明显的亲社会行为, 例如为他人挺身而出(Bergin et al., 2003)。最后, 中国家庭教育促进法的颁布为家庭教育提供了明确的法律框架, 强调了家庭在培养青少年良好心理适应能力方面的责任。为青少年提供更为健康、和谐的家庭环境, 这对他们的社会适应具有积极作用。且前段时间教育部等十三部委发布关于健全家校协同育人意见, 通过促使学校与家庭更紧密地合作, 共同致力于青少年的全面发展。
尽管有这些优势, 但本研究仍存在一些应谨慎考虑的限制与不足。首先, 本研究样本虽然具有较好的统计功效, 但本研究均来自同一学校的学生, 在样本代表性和数量上仍存在一些差距。其次, 虽然基于纵向设计, 但要求青少年回忆他们童年时期的虐待经历。这可能会引入回忆偏差和更多的测量误差, 且未控制T1时间点的社会适应, 因此研究结果可能存在一定偏差。同时, 不能排除反向因果关系的可能性, 即抑郁症状越多的青少年可能更容易回忆起不良的童年经历(Tsehay et al., 2020)。因此, 在未来的研究中使用跨越童年和青春期的前瞻性设计可能有助于建立变量的时间顺序。不仅如此, 在本研究NR3C1多态性rs6198位点的MAF为0.0002, 虽然与北京千人计划组的频率类似, 但相比国外类似研究(Bouma et al., 2011; Rovaris et al., 2013), 其存在一定的差异, 这种情况可能是由于样本选择、地理差异或其他人群特异性因素导致。此外, 本研究将亲子关系视为“近期压力因素”, 以便探究不同环境类型和遗传因素对社会适应的影响。传统上, 亲子关系被视为积极因素, 但将其纳为压力因素可能会引起读者的负面体验。未来的研究可以考虑通过测量不同养育方式, 如温暖、专制等, 来评估亲子关系的影响。最后, 由于未发现G × E × E三者交互效应对青少年自伤行为的影响, 补充研究中仅测量了抑郁症状和亲社会行为, 可能影响研究的全面性, 且在补充研究中我们并未发现G × E × E三者交互效应对亲社会行为等影响, 可能是由于研究范式调查的亲社会行为类型不符合青少年群体, 研究显示青少年在移情关注、换位思考等方面更具优势(Gregory et al., 2009)。
6 结论
综上, 本研究证明HPA轴系统累积遗传变异得分调节青少年的压力敏感性, 具体表现为, HPA轴系统MGPS调节童年期创伤与亲子关系的交互作用对青少年社会适应的影响, 且高遗传敏感性个体对环境刺激的影响机制符合差别易感性模型, 即积极环境中表现出低水平抑郁症状、自伤行为和更高的亲社会行为, 而且消极环境中有着更高的抑郁症状、自伤行为和更低的亲社会行为。结果提示未来需要对差异遗传敏感性的实际影响进行研究, 相关干预与咨询计划可能需要量身定制, 针对不同遗传敏感性的儿童和青少年给予不同的学习计划和奖励方式(Bakermans-Kranenburg & Van Ijzendoorn, 2015)。
附录1. 抑郁症状、亲社会行为和自伤行为指标选择原因与意义
在主实验中, 我们采用了传统的遗传与环境交互研究范式, 结合问卷调查的方式, 以全面评估青少年社会适应的多个方面。通过使用量表, 我们测量了青少年在抑郁症状、亲社会行为和自伤行为等方面的表现。这些指标不仅是遗传与环境研究中经典且常见的测量工具, 还在HPA轴系统的调控下发挥关键作用, 同时也是青少年心理健康的综合反映。
首先, 抑郁症状的评估在研究中具有重要价值, 不仅能够帮助我们深入了解青少年在情感和心理层面的适应状况, 而且作为遗传与环境研究范式的经典因素, 一直以来受到研究者的广泛关注(Lesch, 2004; Mandelli & Serretti, 2013)。这是因为抑郁症状不仅对个体心理健康产生直接影响, 而且在青少年时期的表现可能与日后成年期的心理健康问题有关(Gargano et al., 2018)。青少年是个体心理、生理和社会化方面迅速发展的阶段, 同时也是抑郁出现的高发期(Dunn et al., 2012)。值得注意的是, 根据2022年中国心理健康蓝皮书, 我国青少年抑郁的检出率已经达到了24.6%。这表明抑郁问题在我国青少年中广泛存在, 对他们的心理健康产生了重要影响。另外, 我国卫生健康委员会于2020年9月11日专门发文提出, 将抑郁筛查纳入高中及高等院校健康体检内容, 这反映了对青少年抑郁问题的社会关切;
亲社会行为的测量是研究中的关键组成部分, 其聚焦点在于了解青少年与家庭以及社会之间的积极互动。这一维度不仅是一种积极的心理特征, 还是青少年在社会环境中适应的重要体现。将亲社会行为作为研究对象有助于深入了解青少年在日常生活中与他人的交往方式、合作倾向和社会支持体验(Eberly & Montemayor, 1998; Wentzel et al., 2007)。亲社会行为在研究中具有特殊的意义, 因为它可以被视为一个正面心理指标。通过这一指标的测量, 我们能够验证差别易感性模型, 即在积极的家庭和社会环境中, 青少年可能会表现出更好的亲社会行为, 而在消极的环境中可能表现更差。这种差别易感性模型对于理解遗传与环境交互对青少年社会适应的影响机制具有重要意义;
自伤行为的考察对于研究青少年的心理适应和压力应对方式提供了重要的洞察。通过深入了解青少年如何应对内在和外在压力, 我们可以揭示他们可能面临的困境, 并为制定有针对性的心理干预策略提供依据。此外, 在课题组的一线心理干预与实践中, 我们发现青少年自伤行为普遍存在。几乎所有主动咨询的孩子中都可观察到自伤行为的迹象。这一发现引起了我们对青少年心理健康问题的深刻关注。自伤行为可能是青少年面对压力和情感困扰时的一种非典型应对方式, 其出现可能提示了他们内在困扰的存在。
这些方面的研究不仅提供了对个体心理健康状况的深刻洞察, 还为理解遗传与环境因素如何相互作用以影响这些行为和症状的表现提供了重要线索。
其次, 我们之所以选择抑郁、亲社会和自伤行为作为研究指标, 不仅是因为它们在遗传与环境研究中被广泛应用, 更是因为它们与HPA轴系统密切相关。HPA轴系统作为身体应激响应的一部分, 在情感调控、社会适应等方面发挥着至关重要的作用。这一系统在应对压力和适应环境变化时起着关键的调节作用, 与心理健康紧密相连。大量的研究已经显示, HPA轴与抑郁、亲社会和自伤行为之间存在显著的相关性(Guerry & Hastings, 2011; Endedijk et al., 2019; Klimes-Dougan et al., 2019)。情感障碍, 如抑郁症状, 与HPA轴的活动水平有关, 这反映了身体对心理压力的生理性回应。亲社会行为的表现和社会适应也与HPA轴的稳定性和敏感性密切相关。自伤行为可能与HPA轴的过度激活或不足反应有关, 成为应对心理压力的一种方式。
因此, 我们选择这些指标不仅是因为它们是传统测量工具, 更是因为它们在研究HPA轴与心理健康之间关系时提供了独特的视角。通过对这些指标的深入研究, 我们可以更全面地理解遗传与环境交互如何影响HPA轴的活动, 从而进一步了解这一系统与青少年心理健康之间的复杂关系。这有助于推动对青少年心理适应机制的深入认识, 为未来的预防和干预工作提供更为精准的依据。
参考文献:
Dunn, B. D. (2012). Helping depressed clients reconnect to positive emotion experience: Current insights and future directions. Clinical Psychology & Psychotherapy, 19(4), 326−340.
Eberly, M. B., & Montemayor, R. (1998). Doing good deeds: An examination of adolescent prosocial behavior in the context of parent-adolescent relationships. Journal of Adolescent Research, 13(4), 403−432.
Endedijk, H. M., Nelemans, S. A., Schür, R. R., Boks, M. P., Van Lier, P., Meeus, W.,... Branje, S. (2019). The role of stressful parenting and mineralocorticoid receptor haplotypes on social development during adolescence and young adulthood. Journal of Youth and Adolescence, 48, 1082−1099.
Gargano, L. M., Locke, S., Li, J., & Farfel, M. R. (2018). Behavior problems in adolescence and subsequent mental health in early adulthood: results from the World Trade Center Health Registry Cohort. Pediatric Research, 84(2), 205−209.
Guerry, J. D., & Hastings, P. D. (2011). In search of HPA axis dysregulation in child and adolescent depression. Clinical Child and Family Psychology Review, 14, 135−160.
Klimes-Dougan, B., Begnel, E., Almy, B., Thai, M., Schreiner, M. W., & Cullen, K. R. (2019). Hypothalamic-pituitary- adrenal axis dysregulation in depressed adolescents with non-suicidal self-injury. Psychoneuroendocrinology, 102, 216−224.
Lesch, K. P. (2004). Gene-environment interaction and the genetics of depression. Journal of Psychiatry and Neuroscience, 29(3), 174−184.
Mandelli, L., & Serretti, A. (2013). Gene environment interaction studies in depression and suicidal behavior: an update. Neuroscience & Biobehavioral Reviews, 37(10), 2375−2397.
Wentzel, K. R., Filisetti, L., & Looney, L. (2007). Adolescent prosocial behavior: The role of self-processes and contextual cues. Child Development, 78(3), 895−910.
附录2. 实验设计及操作
本实验采用本公司的imLDRTM多重SNP分型试剂盒对SNP位点分型。
(1)目标SNP位点所在区段先采用多重PCR反应在一个体系中获得扩增。
(2)扩增产物经外切核酸酶及虾碱酶(ExoI/SAP)纯化后用于后续连接酶反应的模板。
(3)在一个连接反应中, 每个位点包含两个5’端等位基因特异探针(其3′末端分别为两个等位基因特异碱基或序列——对插入缺失多态而言)以及紧挨其后的一条3′端位点的荧光标记的特异探针。连接产物通过ABI3730XL的毛细管电泳来区分, 原始数据文件用GeneMapper 4.1软件(Appliedbiosystems)来分析。
具体实验操作步骤:
1)DNA样本取1μl 1% agarose电泳对其样本呢进行质量检查以及浓度估计, 然后根据估计的浓度将样本稀释到工作浓度5-10 ng/μl.
2)多重PCR反应:
a) PCR 条件
反应体系(10 μl)包含1x GC-I buffer (Takara.), 3.0 mM Mg2+, 0.3 mM dNTP, 1 U HotStarTaq polymerase (Qiagen Inc.), 1 µl样本DNA和1 µl多重PCR引物。
3)多重PCR产物纯化
在10 μl PCR产物中加入5U SAP酶和2U Exonuclease I酶, 37℃温浴1小时, 然后75℃灭活15分钟。
b)连接反应
反应体系:10x连接缓冲液1 ul、高温连接酶0.25 ul、5′连接引物混合液(1 µM) 0.4 ul, 3′连接引物混合液(2 µM) 0.4 ul、纯化后多重PCR产物2 ul、ddH2O 6 ul混匀。
4)连接产物上ABI3730XL测序仪
取0.5 μl稀释后的连接产物, 与0.5 μl Liz500 SIZE STANDARD, 9 μl Hi-Di混匀, 95℃变性5分钟后上ABI3730XL测序仪
5) ABI3730XL测序仪上收集的原始数据用GeneMapper 4.1 (AppliedBiosystems, USA)来分析。
附录3. HPA轴系统相关SNP与其功能的关系以及编码方案
SKA2 rs7208505:纺锤体和着丝粒相关复合物亚单位2 (spindle and kinetochore-associated complex subunit 2)编码一种蛋白质, 参与细胞分裂过程中纺锤体和着丝粒的形成。纺锤体和着丝粒是细胞分裂时非常重要的结构, 它们有助于正确分离染色体, 确保新生细胞得到正确的染色体数量。过往研究研究表明, SKA2基因可能与精神健康状况有关(Rice et al., 2008)。rs7208505是位于SKA2 3'非翻译区的一个特异性变体。在一项探究SKA2 rs7208505与自伤关系的研究中, Valenzuela-García等人(2023)发现自杀受害者中rs7208505等位基因A的比例高于健康对照组。不仅如此, Ijabi等人(2019)结果也显示, 相对G/G基因型的孕妇, A/A基因型的孕妇起早产后焦虑风险增加了9.6倍。
NR3C1基因编码核受体激素亲和素1(nuclear receptor subfamily 3 group C member 1, NR3C1), 也被称为葡萄糖皮质激素受体(GR, Glucocorticoid Receptor)。这个基因位于人类染色体5号上, 是一个重要的基因, 参与调节机体对于葡萄糖皮质激素(如皮质醇)的反应。NR3C1基因编码的核受体是一种转录因子, 它在细胞内调节基因的转录, 影响细胞的生长、发育和代谢。葡萄糖皮质激素是一类由肾上腺皮质分泌的激素, 对于机体的代谢、免疫反应和应激反应等方面有重要的调节作用 (Arai et al., 2003)。NR3C1基因的功能异常或变异可能与多种疾病和生理过程有关, 包括内分泌失调、免疫系统异常、心血管疾病等。此外, NR3C1基因也在神经科学和精神病学领域引起关注, 因为它参与调节应激响应, 与精神健康和情绪调节有关。一些研究表明NR3C1基因的变异可能与抑郁症、焦虑症等精神障碍的风险相关。总体而言, NR3C1基因在生理和病理条件下的调控作用使其成为生命科学和医学研究中的一个重要对象 (van Leeuwen et al., 2011)。
NR3C1 rs6198: Rovaris等人(2013)研究发现, NR3C1基因(rs6198 C等位基因)可能与每天吸烟数量和尼古丁依赖性相关, 而NR3C2基因的多态性也可能与吸烟行为有关。此外, 在一项探究NR3CI, NR3C2和童年期创伤交互作用对自杀未遂的研究中, 结果显示rs6198 C等位基因携带者中NR3C1基因过表达, 增加了自杀未遂的风险(Sanabrais-Jiménez et al., 2023)。
NR3C1 rs41423247:根据Pagliaccio等人(2014)在其发表在neuropsychopharmacology使用MGPS方法的研究中, 确定G等位是影响HPA轴的风险等位基因。且相关研究显示携带G等位基因的个体特里尔压力测试下皮质醇增加 (Duncan & Keller, 2011), 且会增加重症抑郁症发作风险 (Sarubin et al., 2017)
NR3C2 基因编码核受体激素亲和素2 (nuclear receptor subfamily 3 group C member 2, NR3C2), 或称为"mineralocorticoid receptor" (矿物质皮质激素受体)基因编码的是矿物质皮质激素受体, 也被称为盐皮质激素受体(MR, Mineralocorticoid Receptor)。这个基因位于人类染色体4号上, 是一个核受体基因, 与糖皮质激素受体(GR, Glucocorticoid Receptor, 由NR3C1基因编码)一同参与调节体内激素的平衡。盐皮质激素受体对于矿物质皮质激素(如醛固酮)的结合和调节在体内维持水电解质平衡、血压稳定等方面发挥着关键作用。此外, 与NR3C1基因相似, NR3C2基因也涉及到应激响应、免疫系统等生理过程。
NR3C2 rs17581262: 有研究发现rs17581262的G等位基因与较小的海马和杏仁核体积, 以及与遭受过儿童期受虐的个体中较高的重症抑郁风险相关(Gerritsen et al., 2017)。
FKBP5: 全称是"FK506 Binding Protein 5", 中文翻译为"FK506结合蛋白5"。FKBP5是一种人体基因, 编码一种蛋白质, 它在细胞内的功能涉及到蛋白质折叠和信号转导的调控。这个基因的名称中, “FKBP”表示蛋白质与一种免疫抑制药物FK506(孤儿草霉素)结合, 而“5”表示这是该家族的第五个成员。FKBP5蛋白在神经系统中发挥重要作用, 特别是在应激和神经适应性方面。它与应激激素(如皮质醇)的信号传导有关, 参与了身体对压力的应对机制。FKBP5基因的变异与一些精神健康障碍, 如抑郁症和创伤后应激障碍(PTSD)等有关。
FKBP5 rs1360780: 根据Pagliaccio等人(2014)在其发表在neuropsychopharmacology使用MGPS方法的研究中, 确定T等位是影响HPA轴的风险等位基因。此外, FKBP5基因的T等位基因(rs1360780)携带者在大脑结构上表现出较小的背侧前部扣带皮层体积以及较高的平均扩散率值, 与非携带者相比, 这可能与应激相关精神疾病如创伤后应激障碍(PTSD)的结构基础有关(Fujii et al., 2014)。
FKBP5 rs9470080: 相比健康对照组, 功能性癫痫发作和重度抑郁症患者的T等位基因显著增加(Asadi‐Pooya et al., 2023), 而且在两个独立的复制样本中也发现了T等位基因退役军人携带者, 遭受儿童期受虐后创伤后应激障碍显著增加 (Watkins et al., 2016)
参考文献:
Arai K, Nakagomi Y, Iketani M, Shimura Y, Amemiya S, Ohyama K et al (2003) Functional polymorphisms in the mineralocorticoid receptor and amirolide-sensitive sodium channel genes in a patient with sporadic pseudohypoaldosteronism. Hum Genet 112(1), 91−97
Duncan LE, Keller MC (2011). A critical review of the first 10 years of candidate gene-by-environment interaction research in psychiatry. Am J Psychiatry 168, 1041-1049.
Fujii, T., Ota, M., Hori, H., Hattori, K., Teraishi, T., Sasayama, D.,... Kunugi, H. (2014). Association between the common functional FKBP5 variant (rs1360780) and brain structure in a non-clinical population. Journal of Psychiatric Research, 58, 96−101.
Gerritsen, L., Milaneschi, Y., Vinkers, C. H., Van Hemert, A. M., Van Velzen, L., Schmaal, L., & Penninx, B. W. (2017). HPA axis genes, and their interaction with childhood maltreatment, are related to cortisol levels and stress- related phenotypes. Neuropsychopharmacology, 42(12), 2446−2455.
Ijabi, J., Moradi-Sardareh, H., Afrisham, R., Seifi, F., & Ijabi, R. (2019). SKA2 gene-A novel biomarker for latent anxiety and preterm birth prediction. European Journal of Obstetrics & Gynecology and Reproductive Biology, 237, 106−112.
Rice, L., Waters, C. E., Eccles, J., Garside, H., Sommer, P., Kay, P.,... Ray, D. W. (2008). Identification and functional analysis of SKA2 interaction with the glucocorticoid receptor. The Journal of Endocrinology, 198(3), 499.
Rovaris, D. L., Mota, N. R., de Azeredo, L. A., Cupertino, R. B., Bertuzzi, G. P., Polina, E. R.,... Bau, C. H. (2013). MR and GR functional SNPs may modulate tobacco smoking susceptibility. Journal of Neural Transmission, 120, 1499−1505.
Sanabrais-Jiménez, M. A., Esquivel-López, A. A., Sotelo- Ramírez, C. E., Aguilar-García, A., Ordoñez-Martínez, B., Jiménez-Pavón, J.,... Camarena, B. (2023). NR3C1 and NR3C2 genes increase the risk of suicide attempt in psychiatric disorder patients with history of childhood trauma. Neuropsychiatric Disease and Treatment, 2561− 2571.
Sarubin, N., Hilbert, S., Naumann, F., Zill, P., Wimmer, A. M., Nothdurfter, C.,... Schüle, C. (2017). The sex-dependent role of the glucocorticoid receptor in depression: variations in the NR3C1 gene are associated with major depressive disorder in women but not in men. European Archives of Psychiatry and Clinical Neuroscience, 267, 123−133.
Valenzuela-García, L. I., Ayala-García, V. M., Ramos-Rosales, D. F., Jacquez-Flores, R. E., Urtiz-Estrada, N., Hernández, E. M. M., & Barraza-Salas, M. (2023). The rs7208505 polymorphism and differential expression of the SKA 2 gene in the prefrontal cortex of suicide victims from the Mexican population. Archives of Suicide Research, 1−12.
van Leeuwen N, Bellingrath S, de Kloet ER, Zitman FG, DeRijk RH, Kudielka BM et al (2011) Human mineralocorticoid receptor (MR) gene haplotypes modulate MR expression and transactivation: Implication for the stress response. Psychoneuroendocrinology 36(5): 699- 709
Watkins, L. E., Han, S., Harpaz-Rotem, I., Mota, N. P., Southwick, S. M., Krystal, J. H.,... Pietrzak, R. H. (2016). FKBP5 polymorphisms, childhood abuse, and PTSD symptoms: Results from the National Health and Resilience in Veterans Study. Psychoneuroendocrinology, 69, 98−105.
附录4. 遗传与MGPS Johnson-Neyman图
附录5. HPA轴系统MGPS和童年期创伤对青少年自伤行为的影响
| 变量 | 自伤行为 | |||
|---|---|---|---|---|
| β | t | p | 95% CI | |
| 童年期创伤 | 0.36 | 10.28*** | < 0.001 | (0.29, 0.43) |
| MGPS | 0.02 | 0.57 | 0.57 | (−0.05, 0.09) |
| G × E | 0.09 | 2.49* | 0.01 | (0.02, 0.15) |
| R2 | 0.14 | |||
| F | 36.62*** | |||
附录6. 单个基因位点与环境的交互作用对青少年社会适应的影响
表1 父子模型
| 变量 | 抑郁症状 | 亲社会行为 | 自伤行为 | |||
|---|---|---|---|---|---|---|
| β | p | β | p | β | p | |
| rs7208505 | 0.005 | 0.92 | −0.10 | 0.06 | 0.07 | 0.19 |
| rs6198 | / | / | / | / | / | / |
| rs41423247 | 0.02 | 0.73 | −0.04 | 0.55 | 0.07 | 0.26 |
| rs17581262 | 0.1003 | 0.64 | 0.08 | 0.20 | 0.07 | 0.28 |
| rs1360780 | 0.10*** | < 0.001 | −0.05 | 0.34 | 0.14* | 0.01 |
| rs9470080 | 0.14** | 0.003 | −0.05 | 0.36 | 0.03 | 0.61 |
表2 母子模型
| 变量 | 抑郁症状 | 亲社会行为 | 自伤行为 | |||
|---|---|---|---|---|---|---|
| β | p | β | p | β | p | |
| rs7208505 | 0.05 | 0.32 | −0.10 | 0.07 | 0.07 | 0.19 |
| rs6198 | / | / | / | / | / | / |
| rs41423247 | 0.02 | 0.75 | −0.02 | 0.77 | 0.07 | 0.26 |
| rs17581262 | 0.10 | 0.09 | 0.004 | 0.96 | 0.07 | 0.28 |
| rs1360780 | 0.08 | 0.12 | −0.10 | 0.07 | 0.14* | 0.01 |
| rs9470080 | 0.06 | 0.19 | −0.08 | 0.12 | 0.03 | 0.61 |
附录7. HPA轴系统基因对青少年适应主效应分析
表1 HPA轴系统单基因对青少年心理适应主效应分析
| 变量 | 抑郁症状 | 亲社会行为 | 自伤行为 | |||
|---|---|---|---|---|---|---|
| β | p | β | p | β | p | |
| rs7208505 | −0.02 | 0.57 | 0.01 | 0.90 | 0.05 | 0.21 |
| rs6198 | 0.03 | 0.51 | 0.03 | 0.35 | −0.02 | 0.64 |
| rs41423247 | −0.04 | 0.30 | 0.09 | 0.02* | −0.03 | 0.43 |
| rs17581262 | 0.02 | 0.67 | 0.05 | 0.17 | −0.002 | 0.95 |
| rs1360780 | −0.04 | 0.33 | 0.004 | 0.92 | 0.03 | 0.52 |
| rs9470080 | −0.05 | 0.16 | 0.05 | 0.23 | −0.01 | 0.75 |
附录8. 独立的HPA轴系统(n-1)MGPSs与环境的交互作用对青少年社会适应的影响
表1 独立的HPA轴系统(n-1)MGPSs与环境的交互作用对青少年社会适应的影响(父子模型)
| 变量 | 抑郁症状 | 亲社会行为 | 自伤行为 | |||
|---|---|---|---|---|---|---|
| β | p | β | p | β | p | |
| -rs7208505 | 0.07** | 0.002 | / | / | 0.05* | 0.03 |
| -rs6198 | 0.05** | 0.003 | / | / | 0.05* | 0.01 |
| -rs41423247 | 0.06** | 0.002 | / | / | 0.05* | 0.02 |
| -rs17581262 | 0.05** | 0.005 | / | / | 0.04* | 0.03 |
| -rs1360780 | 0.06** | 0.003 | / | / | 0.06* | 0.047 |
| -rs9470080 | 0.06** | 0.01 | / | / | 0.09** | 0.002 |
表2 独立的HPA轴系统(n-1)MGPSs与环境的交互作用对青少年社会适应的影响(母子模型)
| 变量 | 抑郁症状 | 亲社会行为 | 自伤行为 | |||
|---|---|---|---|---|---|---|
| β | p | β | p | β | p | |
| -rs7208505 | 0.04* | 0.04 | −0.04 | 0.12 | 0.05* | 0.03 |
| -rs6198 | 0.04* | 0.03 | −0.04* | 0.047 | 0.05* | 0.01 |
| -rs41423247 | 0.05* | 0.02 | −0.04* | 0.05 | 0.05* | 0.02 |
| -rs17581262 | 0.03 | 0.12 | −0.05* | 0.05 | 0.04* | 0.03 |
| -rs1360780 | 0.06* | 0.03 | −0.06* | 0.05 | 0.06* | 0.047 |
| -rs9470080 | 0.06* | 0.03 | −0.06* | 0.048 | 0.09** | 0.002 |
附录9. 内部一致性分析
表2 青少年社会适应的G × E × E检验 (父子模型sample 1)
| 变量 | 抑郁症状 | 亲社会行为 | 自伤行为 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β | t | p | 95% CI | β | t | p | 95% CI | β | t | p | 95% CI | |
| 亲子关系 | −0.31 | −5.77*** | < 0.001 | (−0.41, −0.04) | 0.04 | 0.78 | 0.44 | (−0.07, 0.16) | / | / | / | / |
| 童年期创伤 | 0.27 | 4.49*** | < 0.001 | (0.15, 0.39) | −0.11 | −1.59 | 0.11 | (−0.24, 0.02) | 0.33 | 6.56*** | < 0.001 | (0.23, 0.43) |
| MGPS | −0.03 | −0.53 | 0.60 | (−0.12, 0.07) | 0.04 | 0.71 | 0.48 | (−0.07, 0.14) | 0.01 | 0.30 | 0.77 | (−0.08, 0.11) |
| E1 × E2 | −0.05 | −1.06 | 0.29 | (−0.14, 0.04) | −0.03 | −0.58 | 0.57 | (−0.13, 0.07) | 0.06 | −1.32 | 0.16 | (−0.03, 0.15) |
| E1 × G | −0.03 | −0.68 | 0.50 | (−0.12, 0.06) | −0.03 | −0.64 | 0.52 | (−0.13, 0.07) | / | / | / | / |
| E2 × G | 0.03 | 0.64 | 0.52 | (−0.07, 0.14) | −0.01 | −0.08 | 0.93 | (−0.12, 0.11) | / | / | / | / |
| E1 × E2 × G | −0.01 | −0.12 | 0.90 | (−0.09, 0.08) | −0.04 | −0.79 | 0.43 | (−0.12, 0.05) | / | / | / | / |
| R2 | 0.24 | 0.02 | 0.12 | |||||||||
| F | 15.23*** | 1.04 | 15.17*** | |||||||||
注:E1为亲子关系; E2为童年期创伤; G为MGPS, 加粗字体表示显著。
表3 青少年社会适应的G × E × E检验 (父子模型sample 2)
| 变量 | 抑郁症状 | 亲社会行为 | 自伤行为 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β | t | p | 95% CI | β | t | p | 95% CI | β | t | p | 95% CI | |
| 亲子关系 | −0.36 | −7.31*** | < 0.001 | (−0.46, −0.26) | 0.16 | 3.09** | 0.002 | (0.06, 0.27) | / | / | / | / |
| 童年期创伤 | 0.16 | 3.12*** | 0.002 | (0.06, 0.26) | −0.18 | −3.47** | 0.001 | (−0.29, −0.08) | 0.41 | 8.05*** | < 0.001 | (0.31, 0.50) |
| MGPS | 0.07 | 1.38 | 0.17 | (−0.03, 0.18) | −0.05 | −0.97 | 0.33 | (−0.16, 0.06) | 0.02 | 0.45 | 0.65 | (−0.08, 0.13) |
| E1 × E2 | 0.04 | 0.71 | 0.48 | (−0.07, 0.14) | −0.16 | −2.78** | 0.01 | (−0.27, −0.05) | 0.13 | 2.47* | 0.01 | (0.03, 0.23) |
| E1 × G | −0.07 | −1.40 | 0.16 | (−0.17, 0.03) | 0.08 | 1.54 | 0.12 | (−0.02, 0.19) | / | / | / | / |
| E2 × G | 0.04 | 0.69 | 0.49 | (−0.07, 0.15) | −0.01 | −0.12 | 0.91 | (−0.12, 0.11) | / | / | / | / |
| E1 × E2 × G | 0.09 | 1.87 | 0.07 | (−0.03, 0.18) | −0.04 | −0.74 | 0.46 | (−0.15, 0.07) | / | / | / | / |
| R2 | 0.19 | 0.10 | 0.16 | |||||||||
| F | 11.51*** | 5.26*** | 22.21*** | |||||||||
注:E1为亲子关系; E2为童年期创伤; G为MGPS, 加粗字体表示显著。
表4 青少年社会适应的G × E × E检验 (母子模型sample 1)
| 变量 | 抑郁症状 | 亲社会行为 | 自伤行为 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β | t | p | 95% CI | β | t | p | 95% CI | β | t | p | 95% CI | |
| 亲子关系 | −0.24 | −4.48*** | < 0.001 | (−0.34, −0.13) | 0.11 | 1.95 | 0.05 | (−0.001, 0.22) | / | / | / | / |
| 童年期创伤 | 0.37 | 6.66*** | < 0.001 | (0.26, 0.47) | −0.09 | −1.54 | 0.13 | (−0.20, 0.02) | 0.33 | 6.56*** | < 0.001 | (0.23, 0.43) |
| MGPS | −0.03 | −0.61 | 0.54 | (−0.13, 0.07) | 0.02 | 0.41 | 0.68 | (−0.08, 0.12) | 0.01 | 0.30 | 0.77 | (−0.08, 0.11) |
| E1 × E2 | 0.03 | 0.67 | 0.50 | (−0.06, 0.12) | −0.02 | −0.49 | 0.62 | (−0.12, 0.07) | 0.06 | −1.32 | 0.16 | (−0.03, 0.15) |
| E1 × G | −0.08 | −1.82 | 0.07 | (−0.17, 0.01) | −0.08 | −1.90 | 0.06 | (−0.18, 0.003) | / | / | / | / |
| E2 × G | −0.01 | −0.14 | 0.89 | (−0.11, 0.09) | −0.03 | −0.52 | 0.60 | (−0.13, 0.08) | / | / | / | / |
| E1 × E2 × G | −0.01 | −0.26 | 0.79 | (−0.09, 0.07) | −0.08 | −1.86 | 0.06 | (−0.15, 0.001) | / | / | / | / |
| R2 | 0.21 | 0.05 | 0.12 | |||||||||
| F | 13.09*** | 2.29* | 15.17*** | |||||||||
注:E1为亲子关系; E2为童年期创伤; G为MGPS, 加粗字体表示显著。
表5 青少年社会适应的G × E × E检验 (母子模型sample 2)
| 变量 | 抑郁症状 | 亲社会行为 | 自伤行为 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β | t | p | 95% CI | β | t | p | 95% CI | β | t | p | 95% CI | |
| 亲子关系 | −0.19 | −4.16*** | <0.001 | (−0.28, −0.10) | 0.15 | 2.83** | 0.005 | (0.05, 0.25) | / | / | / | / |
| 童年期创伤 | 0.34 | 7.05*** | <0.001 | (0.24, 0.43) | −0.19 | −2.83*** | <0.001 | (−0.29, −0.08) | 0.41 | 8.05*** | <0.001 | (0.31, 0.50) |
| MGPS | −0.05 | −0.99 | 0.32 | (−0.10, 0.09) | −0.08 | −1.35 | 0.18 | (−0.19, 0.03) | 0.02 | 0.45 | 0.65 | (−0.08, 0.13) |
| E1 × E2 | −0.01 | −0.13 | 0.90 | (−0.10, 0.09) | −0.15 | −2.75** | 0.01 | (−0.26, −0.04) | 0.13 | 2.47* | 0.01 | (0.03, 0.23) |
| E1 × G | −0.03 | −0.55 | 0.58 | (−0.13, 0.07) | 0.04 | 0.77 | 0.44 | (−0.07, 0.16) | / | / | / | / |
| E2 × G | 0.01 | 0.25 | 0.80 | (−0.09, 0.12) | −0.004 | −0.07 | 0.94 | (−0.12, 0.12) | / | / | / | / |
| E1 × E2 × G | 0.09 | −1.83 | 0.07 | (−0.16, 0.04) | −0.06 | −1.00 | 0.32 | (−0.18, 0.06) | / | / | / | / |
| R2 | 0.20 | 0.09 | 0.16 | |||||||||
| F | 12.36*** | 4.31*** | 22.21*** | |||||||||
附录10
1 情绪Sternberg任务
使用E-prime 2.0呈现刺激材料。程序示意图见图1, 屏幕中央首先呈现注视点“+”并持续300 ms, 随后呈现300 ms的空屏, 接着目标刺激会出现在屏幕中央, 被试在保证准确的情况下尽可能快地对目标的颜色进行反应, 直到被试做出反应或达到反应上限2000 ms, 目标消失, 最后再呈现300 ms的空屏进入下一试次。在本任务中, 目标刺激是红色、绿色或蓝色词。目标刺激若为红色词, 要求被试用左手食指按“F”键反应, 若为绿色词, 要求被试用右手食指按“J”键反应, 若为蓝色词, 要求被试用右手中指按“K”键反应。练习实验由10个试次组成, 正式实验由120个试次组成, 红、绿和蓝色分别包括40个试次, 其中, 每种颜色内, 积极、消极、中性和抑郁词各10个。本实验的测试指标为被试判断的反应时和正确率。
最初的或经典的Stroop任务(Stroop, 1935)涉及用不协调的彩色墨水呈现颜色词(例如, 用蓝色墨水打印的“红色”一词)。要求参与者尽快说出每个刺激词的墨水颜色, 同时尝试忽略该词的含义。这种为了命名墨水颜色而抑制单词含义的尝试已被可靠地证明, 与颜色命名一致刺激(例如, 用红色墨水打印的单词“红色”)相比, 会导致更长的响应延迟, 这种现象已被证实。称为Stroop效应(MacLeod, 1991)。Stroop研究中的因变量要么是反应延迟, 即刺激开始和参与者反应之间的时间(以毫秒为单位), 要么是干扰指数, 通常测量为不一致和一致刺激之间的反应延迟差异, 或者不一致和一致刺激之间的反应延迟差异。Stroop任务被视为精神病理学研究中的有用工具, 用于检查所调查疾病的认知过程(Williams et al., 1996)。
后Stroop任务被修改为“Stroop”或“Modified Stroop”, 用于精神病理学研究, 通过将单词刺激的内容从颜色(例如红色、蓝色、绿色)更改为与疾病相关的充满情感的主题在调查中。例如, Stroop在抑郁症研究中使用了悲伤、沮丧和疲倦等词语(MacLeod, 1991)。与中性词相比, 与障碍相关词的延迟命名相关的相对反应潜伏期被认为反映了对障碍相关刺激的注意力偏差(Gotlib et al., 1996)。同样, 与对照组相比, 有障碍的参与者在Stroop的表现较慢, 被认为反映了特定于障碍的注意力偏差。
情绪Stroop任务的修改是为了更好地探究与抑郁症相关的认知偏差。在这一任务中, 单词刺激的内容被改变为与抑郁症相关的情感主题, 如悲伤、沮丧和疲倦, 而不是颜色。参与者被要求尽快报告每个刺激词的墨水颜色, 同时忽略该词的含义。这种修改使研究者能够更好地了解抑郁症患者在面对与其疾病相关的情感信息时的认知反应。研究使用情绪Stroop任务时, 主要关注的因变量包括反应延迟(反应时)和干扰指数(准确率)。反应延迟是指刺激呈现开始和参与者反应之间的时间, 以毫秒为单位。而干扰指数通常测量为不一致和一致刺激之间的反应延迟差异。在情绪Stroop任务中, 与抑郁症相关的词汇的延迟命名与相对反应潜伏期相关, 这被认为反映了对与疾病相关刺激的注意力偏差(Dalgleish & Watts, 1990; Mogg & Bradley, 2005)。抑郁症患者在情感Stroop任务中通常表现出比对照组更慢的反应(Segal, 2014)。这种延迟被解释为对与抑郁症相关的情感信息的注意力偏差。这与Beck (1979)的抑郁认知理论(cognitive theory of depression)和Bower (1981)的情感网络理论(network theory of emotion)相一致。Beck的模型认为, 早期的生活经历形成了关于自我、世界和未来的负面图式, 这导致了信息处理的偏见。而Bower的情感网络理论强调情感与认知之间的联系, 预测个体更可能关注与其感受一致的材料。此外, 情绪Stroop任务的研究结果还可以通过比较抑郁症样本和对照样本在不同刺激条件下的表现来获得更深层次的理解, 这可能包括与抑郁症特异刺激、消极刺激、积极刺激和中性刺激相关的表现比较(Clark et al., 2000; Segal & Swallow, 1994)。
总的来说, 情绪Stroop任务被认为是一种有用的工具, 用于测量抑郁症患者在处理与其疾病相关的情感信息时的认知表现, 特别是在注意力偏差方面。
再次感谢您的审阅和指导, 通过您的宝贵意见使论文更为完善!
(1) 准确率:
意义:准确率指的是被试在完成任务时正确识别词语颜色的比例。它反映了个体在面对情绪冲突时的执行准确性。较高的准确率通常表明被试更有效地应对任务的认知冲突, 能够准确地识别词语的颜色而不受到情绪干扰。
解释:如果被试在情绪Stroop任务中表现出较高的准确率, 这可能表示其在处理情绪信息时能够保持较好的认知控制和注意力分配。高准确率可能与更强大的认知控制和抑制情感干扰的能力相关。
(2)反应时
意义:反应时是指被试完成任务所需的时间, 即识别词语颜色的反应速度。在情绪Stroop任务中, 较长的反应时通常被视为对情绪冲突的一种指示, 因为被试可能受到了情感信息的干扰, 导致执行任务的速度减慢。
解释:较短的反应时可能表明被试能够更迅速、有效地调整注意力, 克服情感冲突。相反, 较长的反应时可能暗示被试对情感信息的敏感性较高, 需要更多时间来应对冲突。
在实际研究中, 研究者通常结合准确率和反应时来综合评估个体的认知控制水平和情绪调节能力。这种任务在研究情绪和认知之间的关系以及情绪障碍中的认知特征方面具有广泛的应用。
参考文献:
Beck, A. T. (Ed.). (1979). Cognitive therapy of depression. Guilford press.
Bower, G. H. (1981). Mood and memory. American Psychologist, 36(2), 129.
Clark, D. A., Beck, A. T., Alford, B. A., Bieling, P. J., & Segal, Z. V. (2000). Scientific foundations of cognitive theory and therapy of depression.
Dalgleish, T., & Watts, F. N. (1990). Biases of attention and memory in disorders of anxiety and depression. Clinical Psychology Review, 10(5), 589−604.
Gotlib, I. H., & Gilboa, E. (2014). Cognitive interference in depression. Cognitive interference: Theories, methods, and findings, 347.
MacLeod, C. M. (1991). Half a century of research on the Stroop effect: an integrative review. Psychological Bulletin, 109(2), 163.
Mogg, K., & Bradley, B. P. (2005). Attentional bias in generalized anxiety disorder versus depressive disorder. Cognitive Therapy and Research, 29, 29−45.
Segal, Z. V. (2014). Cognitive interference in depressive and anxiety-based disorders. In Cognitive Interference (pp. 325−346). Routledge.
Segal, Z. V., & Swallow, S. R. (1994). Cognitive assessment of unipolar depression: Measuring products, processes and structures. Behaviour Research and Therapy, 32(1), 147−158.
Stroop, J. R. (1935). Studies of interference in serial verbal reactions. Journal of Experimental Psychology, 18(6), 643.
Williams, J. M. G., Mathews, A., & MacLeod, C. (1996). The emotional Stroop task and psychopathology. Psychological Bulletin, 120(1), 3.
2 独裁者游戏范式
参照国内外研究, 本研究将采用独裁者博弈任务来测量个体的合作行为水平(陈思静 等, 2015; Raihani & Mcauliffe, 2014)。独裁者游戏由E-Prime 2.0程序编制。首先, 在屏幕上呈现任务要求, 告知被试将与在另一个房间的实验者匿名在线共同分配一笔金钱, 共10个代币(每个代币可兑换价值3元的礼品)。被试需要进行8次分配任务, 每次接受者都不同。电脑会在两位玩家之间随机分配角色A或者B。为了避免被试受到词语的暗示, 使角色A指代分配者, 角色B指代接受者。角色A有权以任何方式分配这笔金钱, 角色B只能接受分配, 实验报酬与分配数额挂钩。角色B最终知道分配方案, 但二者互不认识, 以后不会再见面。在正式分配环节, 所有被试均被告知已被随机安排为A分配者。分配者的任务是在自己和接受者之间分配10个代币。实验将以被试给对方正式分配的代币数作为亲社会行为。
为了营造真实的在线博弈情境, 实验对接受者设置了基本信息, 包括性别, 年龄, 职业, 共创造了八个接受者, 如第一个接受者编号253, 年龄24岁, 职业是在校大学生, 第二个接受者则是编号140, 年龄23岁, 职业是在校大学生。实验将对八个接受者信息在被试间实施平衡化处理。
流程:告知被试“下面将进行一个在线分钱的小游戏, 由您和在另一个实验室的参加者共同分得一笔金钱(10个代币)。游戏中有两种角色, A分配者和B接受者。分配者有权以任何方式分配这笔钱, 接受者只能接受, 无权拒绝。在游戏中, 电脑程序会在您与另一个参加者之间随机分配角色。你们互不认识, 以后也不会相见, 分配结果将以匿名的方式呈现给对方, 并且最终的实验报酬将以分配的金额挂钩。获得的金额越大, 最终报酬就越多。”阅读完毕之后, 计算机呈现:“现在系统开始指定角色A和B, A有权提出分配, B无法拒绝该分配方
案。”为了更加真实, 计算机屏幕会呈现挑选界面, 并告知被试为分配者, 并说明:“恭喜您成为角色 A, 您可以根据自己的喜好来分配10个代币, 分配所获的收益将与最终实验报酬挂钩。”最后, 进入正式分配环节, 告知被试:“请在下面方框内输入给对方分配的金额数目。”以测量被试合作行为。
附录11
表1 青少年亲社会行为的G × E × E检验
| 变量 | 总分 | I1 | I2 | 变量 | 总分 | I1 | I2 |
|---|---|---|---|---|---|---|---|
| F | p | η2p | F | p | η2p | ||
| 父子关系 | 0.22 | 0.64 | 0.002 | 母子关系 | 0.87 | 0.35 | 0.007 |
| 童年期创伤 | 0.05 | 0.82 | 0.000 | 童年期创伤 | 0.003 | 0.96 | 0.000 |
| MGPS | 0.15 | 0.71 | 0.001 | MGPS | 0.09 | 0.77 | 0.001 |
| E1 × E2 | 0.001 | 0.97 | 0.000 | E1 × E2 | 1.72 | 0.19 | 0.014 |
| E1 × G | 0.63 | 0.43 | 0.005 | E1 × G | 0.02 | 0.89 | 0.000 |
| E2 × G | 0.98 | 0.33 | 0.008 | E2 × G | 0.95 | 0.33 | 0.008 |
| E1 × E2 × G | 0.77 | 0.38 | 0.006 | E1 × E2 × G | 0.002 | 0.97 | 0.000 |
参考文献
Screening for depression in well older adults: Evaluation of a short form of the CES-D
We derived and tested a short form of the Center for Epidemiologic Studies Depression Scale (CES-D) for reliability and validity among a sample of well older adults in a large Health Maintenance Organization. The 10-item screening questionnaire, the CESD-10, showed good predictive accuracy when compared to the full-length 20-item version of the CES-D (kappa =.97, P <.001). Cutoff scores for depressive symptoms were > or = 16 for the full-length questionnaire and > or = 10 for the 10-item version. We discuss other potential cutoff values. The CESD-10 showed an expected positive correlation with poorer health status scores (r =.37) and a strong negative correlation with positive affect (r = -.63). Retest correlations for the CESD-10 were comparable to those in other studies (r =.71). We administered the CESD-10 again after 12 months, and scores were stable with strong correlation of r =.59.
those with epilepsy
DOI:10.1007/s13760-023-02365-0
URL
PMID:37642895
[本文引用: 1]

We aimed to investigate the rates of positive screening for depression, anxiety, stress, and suicide risk in adults with seizures [i.e., well-matched groups of patients with focal epilepsy vs. idiopathic generalized epilepsy (IGE) vs. functional seizures (FS)].This was a cross sectional study. Patients, 19-55 years of age, with a diagnosis of IGE, focal epilepsy or FS were investigated at the outpatient epilepsy clinic at Shiraz University of Medical Sciences, Shiraz, Iran, from September 2022 until January 2023 and during their follow-up visits. We used the validated Farsi version of DASS-21 (Depression-Anxiety-Stress Scale) to investigate and screen for depression, anxiety, and stress in these patients. We also used the Beck Scale for Suicide Ideation (BSSI).Forty patients with focal epilepsy, 40 persons with IGE, and 40 individuals with FS were included. Depression and anxiety were more prevalent among patients with FS compared with those with epilepsy. The rate of stress among patients with FS was not significantly different compared with that in patients with epilepsy. The suicide risks were not significantly different between the groups either.Patients with FS are at high risk for psychiatric comorbidities that is comparable or even worse than that in patients with epilepsy. Specific validated scales to screen for psychiatric comorbidities and suicide risk should be integral components of the evaluation and treatment of all patients with seizures.© 2023. The Author(s) under exclusive licence to Belgian Neurological Society.
Maltreatment, MAOA, and delinquency: Sex differences in gene-environment interaction in a large population-based cohort of adolescents
DOI:10.1007/s10519-010-9356-y
PMID:20734127
[本文引用: 1]
The present study investigated a possible interaction between a functional polymorphism in the MAOA gene promoter (MAOA-VNTR) and childhood maltreatment in the prediction of adolescent male and female delinquency. A cohort of 1,825 high school students, 17-18 years old, completed an anonymous questionnaire during class hours which included questions on childhood maltreatment, sexual abuse, and delinquency. Saliva samples were collected for DNA isolation, and analyzed for the MAOA-VNTR polymorphism. Self-reported maltreatment was a strong risk factor for adolescent delinquent behavior. The MAOA genotype also showed a significant main effect when controlled for maltreatment. Boys with a short variant and girls with one or two long variants of the polymorphism showed a higher risk for delinquency when exposed to maltreatment. Our results confirm previous findings of an interaction between the MAOA-VNTR polymorphism and self-reported maltreatment. Results for boys and girls differ according to MAOA-VNTR genotype and direction of phenotypic expression.
Gene-environment interaction and psychiatric disorders: Review and future directions
Genetic markers of the stress generation model: A systematic review
The hidden efficacy of interventions: Gene× environment experiments from a differential susceptibility perspective
DOI:10.1146/annurev-psych-010814-015407
PMID:25148854
[本文引用: 1]
The efficacy of interventions might be underestimated or even go undetected as a main effect when it is hidden in gene-by-environment (G×E) interactions. This review moves beyond the problems thwarting correlational G×E research to propose genetic differential susceptibility experiments. G×E experiments can test the bright side as well as the dark side of the moderating role of genotypes traditionally considered to represent vulnerability to negative conditions. The differential susceptibility model predicts that carriers of these risk genotypes profit most from interventions changing the environment for the better. The evolutionary background of G×E and differential susceptibility is discussed, and statistical methods for the analysis of differential susceptibility (versus diathesis stress) are reviewed. Then, based on results from 22 randomized G×E experiments, meta-analytic evidence for the differential susceptibility model is presented. Intervention effects are much stronger in the susceptible genotypes than in the nonsusceptible genotypes. The final sections suggest possibilities to broaden the G component in the G×E equation by including genetic pathways, and to broaden the E component by including methylation level and gene expression as promising ways to probe the concept of the environment more deeply and address the perennial issue of what works for whom.
Methylation at SLC6A4 is linked to family history of child abuse: An examination of the Iowa Adoptee sample
For better and for worse: Differential susceptibility to environmental influences
Beyond risk, resilience, and dysregulation: Phenotypic plasticity and human development
How can I deal with missing data in my study?
Prosocial behaviours of young adolescents: A focus group study
This study investigated young adolescents' perceptions of their peers' prosocial behaviours. In eight focus groups, 53 11- to 13-year olds described specific prosocial acts of their peers. Results suggest that traditional research has not addressed the diversity of prosocial behaviours that youth enact, nor emphasized behaviours that are salient to young adolescents. Such behaviours included standing up for others, encouraging others, helping others develop skills, including others who are left out, and being humorous. Facilitating emotional regulation of others emerged as an important component of prosocial behaviour. These data can help guide future research on prosocial development to include a broader array of authentic behaviours of young adolescents.
No associations between single nucleotide polymorphisms in corticoid receptor genes and heart rate and cortisol responses to a standardized social stress test in adolescents: The TRAILS study
DOI:10.1007/s10519-010-9385-6
PMID:20680430
[本文引用: 1]
Previously, sequence variation in the glucocorticoid (GR) and mineralocorticoid (MR) receptor genes (NR3C1 and NR3C2, respectively) have been found to be associated with physiological stress responses to social stress tests in small samples of adult men and oral contraceptives (OC) using women. Associations between single nucleotide polymorphisms (SNPs) in the GR (23EK-rs6190, 9beta-rs6198, BclI-rs4142324) and the MR gene (I180V-rs5522 and -2G/C (rs2070951) with cortisol and heart rate responses to a performance-related social stress task (public speaking and mental arithmetic) were examined in a large sample (n = 553) of adolescents (15-17 years). To make comparisons with previous findings, associations were tested in boys (n = 277), free-cycling (FC) girls (n = 183) and OC users (n = 93). None of the previously reported associations in adults could be replicated in this large adolescent sample. Explanations for non-replication are discussed.
Traumatic stress: effects on the brain
Brain areas implicated in the stress response include the amygdala, hippocampus, and prefrontal cortex. Traumatic stress can be associated with lasting changes in these brain areas. Traumatic stress is associated with increased cortisol and norepinephrine responses to subsequent stressors. Antidepressants have effects on the hippocampus that counteract the effects of stress. Findings from animal studies have been extended to patients with post-traumatic stress disorder (PTSD) showing smaller hippocampal and anterior cingulate volumes, increased amygdala function, and decreased medial prefrontal/anterior cingulate function. In addition, patients with PTSD show increased cortisol and norepinephrine responses to stress. Treatments that are efficacious for PTSD show a promotion of neurogenesis in animal studies, as well as promotion of memory and increased hippocampal volume in PTSD.
Caught between parents: Adolescents' experience in divorced homes
This study examined adolescents' feelings of being caught between parents to see whether this construct helps to explain (1) variability in their postdivorce adjustment and (2) associations between family/child characteristics and adolescent adjustment. Adolescents 10 to 18 years old (N = 522) were interviewed by telephone 4 1/2 years after their parents' separation. Feeling caught between parents was related to high parental conflict and hostility and low parental cooperation. Being close to both parents was associated with low feelings of being caught. The relation between time spent with each parent and feeling caught depended on the coparenting relationship. Adolescents in dual residence were especially likely to feel caught when parents were in high conflict, and especially unlikely to feel caught when parents cooperated. Feeling caught was related to poor adjustment outcomes. Parental conflict was only related to adjustment outcomes indirectly, through adolescents' feelings of being caught.
Genetic aetiology of self-harm ideation and behaviour
DOI:10.1038/s41598-020-66737-9
URL
PMID:32546850
[本文引用: 1]
Family studies have identified a heritable component to self-harm that is partially independent from comorbid psychiatric disorders. However, the genetic aetiology of broad sense (non-suicidal and suicidal) self-harm has not been characterised on the molecular level. In addition, controversy exists about the degree to which suicidal and non-suicidal self-harm share a common genetic aetiology. In the present study, we conduct genome-wide association studies (GWAS) on lifetime self-harm ideation and self-harm behaviour (i.e. any lifetime self-harm act regardless of suicidal intent) using data from the UK Biobank (n > 156,000). We also perform genome wide gene-based tests and characterize the SNP heritability and genetic correlations between these traits. Finally, we test whether polygenic risk scores (PRS) for self-harm ideation and self-harm behaviour predict suicide attempt, suicide thoughts and non-suicidal self-harm (NSSH) in an independent target sample of 8,703 Australian adults. Our GWAS results identified one genome-wide significant locus associated with each of the two phenotypes. SNP heritability (h) estimates were ~10%, and both traits were highly genetically correlated (LDSC r> 0.8). Gene-based tests identified seven genes associated with self-harm ideation and four with self-harm behaviour. Furthermore, in the target sample, PRS for self-harm ideation were significantly associated with suicide thoughts and NSSH, and PRS for self-harm behaviour predicted suicide thoughts and suicide attempt. Follow up regressions identified a shared genetic aetiology between NSSH and suicide thoughts, and between suicide thoughts and suicide attempt. Evidence for shared genetic aetiology between NSSH and suicide attempt was not statistically significant.
Hypothalamic- pituitary-adrenal axis multilocus genetic variation, childhood parenting and adolescent anxiety symptoms: Evidence of cumulative polygenic plasticity
The longitudinal relations among dimensions of parenting styles, sympathy, prosocial moral reasoning, and prosocial behaviors
The development of a measure of prosocial behaviors for late adolescents
The strengthening starts at home: Parent-child relationships, psychological capital, and academic performance - A longitudinal mediation analysis
Aggressive behavior and psychiatric inpatients: A narrative review of the literature with a focus on the European experience
DOI:10.1007/s11920-021-01233-z
URL
PMID:33825996
[本文引用: 1]
We summarized peer-reviewed literature on aggressive episodes perpetrated by adult patients admitted to general hospital units, especially psychiatry or emergency services. We examined the main factors associated with aggressive behaviors in the hospital setting, with a special focus on the European experience.A number of variables, including individual, historical, and contextual variables, are significant risk factors for aggression among hospitalized people. Drug abuse can be considered a trans-dimensional variable which deserves particular attention. Although mental health disorders represent a significant component in the risk of aggression, there are many factors including drug abuse, past history of physically aggressive behavior, childhood abuse, social and cultural patterns, relational factors, and contextual variables that can increase the risk of overt aggressive behavior in the general hospital. This review highlights the need to undertake initiatives aimed to enhance understanding, prevention, and management of violence in general hospital settings across Europe.
Social adjustment, social performance, and social skills: A tri-component model of social competence
The influence of third-party punishment on cooperation: An explanation of social norm activation
DOI:10.3724/SP.J.1041.2015.00389
[本文引用: 1]
<p>Third-party punishment (TPP) plays an important role in both improving cooperation and maintaining social norms. However, Cognitive Evaluation Theory suggests that TPP may also negatively affect cooperation, because TPP reduces the internal motivation of cooperative behaviors. Therefore, the influence of TPP on cooperation may have two different manifestations depending on the specific kind of activated social norms — descriptive norms (what most people actually do) or injunctive norms (what people should do). This study used two experiments to examine the influence of TPP on cooperation. Experiment 1 analyzed the effects of the two different social norms on cooperation without TPP. The subjects (120 university students) participated in a two-round Dictator Game, which used a 2 (high/low descriptive norms) by 2 (high/low injunctive norms) between-subjects design. Experiment 2 (with 300 university students) examined the influence of different TPP frequencies on cooperation. The subjects participated in a four-round Dictator Game with a third-party member who could punish both the dictator and the receiver in Round 2 and 3. In Round 3, the subjects were informed the frequency of TPP (a between-subjects factor), which were controlled by the experimenter on 10 levels ranging from 0% to 90%. The result showed that descriptive norms had a more significant influence in comparison to injunctive norms, and there was a significant interaction between the two types of norms. Descriptive norms played a more important role on cooperation when there was no punisher, whereas injunctive norms' effect on cooperation was stronger when there was a punisher. The results also implied that a low frequency of TPP could successfully increase the level of cooperation, even when the punishment sanction was removed. We also found that higher frequency of TPP reduced the internal motivation on cooperation. An explanation of these effects was that TPP could not only remind subjects of the injunctive norms but also the existence of norm violation. When the perception of norm violation increased with higher frequency of TPP, the perception of descriptive norms decreases and so do cooperative behaviors.</p>
第三方惩罚对合作行为的影响: 基于社会规范激活的解释
Children's social competence in cultural context
Social initiative and behavioral control represent two major dimensions of children's social competence. Cultural norms and values with respect to these dimensions may affect the exhibition, meaning, and development of specific social behaviors such as sociability, shyness-inhibition, cooperation-compliance, and aggression-defiance, as well as the quality and function of social relationships. The culturally guided social interaction processes including evaluations and responses likely serve as an important mediator of cultural influence on children's social behaviors, relationships, and developmental patterns. In this article, we review research on children's social functioning and peer relationships in different cultures from an integrative contextual-developmental perspective. We also review research on the implications of the macro-level social and cultural changes that are happening in many societies for socialization and development of social competence.
Research on the difference between network survey and traditional paper survey
网络调查和传统纸质调查的差异性研究
Gene× Environment interaction and resilience: Effects of child maltreatment and serotonin, corticotropin releasing hormone, dopamine, and oxytocin genes
DOI:10.1017/S0954579412000077
PMID:22559122
[本文引用: 1]
In this investigation, gene-environment interaction effects in predicting resilience in adaptive functioning among maltreated and nonmaltreated low-income children (N = 595) were examined. A multicomponent index of resilient functioning was derived and levels of resilient functioning were identified. Variants in four genes (serotonin transporter linked polymorphic region, corticotropin releasing hormone receptor 1, dopamine receptor D4-521C/T, and oxytocin receptor) were investigated. In a series of analyses of covariance, child maltreatment demonstrated a strong negative main effect on children's resilient functioning, whereas no main effects for any of the genotypes of the respective genes were found. However, gene-environment interactions involving genotypes of each of the respective genes and maltreatment status were obtained. For each respective gene, among children with a specific genotype, the relative advantage in resilient functioning of nonmaltreated compared to maltreated children was stronger than was the case for nonmaltreated and maltreated children with other genotypes of the respective gene. Across the four genes, a composite of the genotypes that more strongly differentiated resilient functioning between nonmaltreated and maltreated children provided further evidence of genetic variations influencing resilient functioning in nonmaltreated children, whereas genetic variation had a negligible effect on promoting resilience among maltreated children. Additional effects were observed for children based on the number of subtypes of maltreatment children experienced, as well as for abuse and neglect subgroups. Finally, maltreated and nonmaltreated children with high levels of resilience differed in their average number of differentiating genotypes. These results suggest that differential resilient outcomes are based on the interaction between genes and developmental experiences.
The adaptive calibration model of stress responsivity
Candidate gene- environment interaction research: Reflections and recommendations
A critical review of the first 10 years of candidate gene-by-environment interaction research in psychiatry
DOI:10.1176/appi.ajp.2011.11020191
PMID:21890791
[本文引用: 1]
Gene-by-environment interaction (G×E) studies in psychiatry have typically been conducted using a candidate G×E (cG×E) approach, analogous to the candidate gene association approach used to test genetic main effects. Such cG×E research has received widespread attention and acclaim, yet cG×E findings remain controversial. The authors examined whether the many positive cG×E findings reported in the psychiatric literature were robust or if, in aggregate, cG×E findings were consistent with the existence of publication bias, low statistical power, and a high false discovery rate.The authors conducted analyses on data extracted from all published studies (103 studies) from the first decade (2000-2009) of cG×E research in psychiatry.Ninety-six percent of novel cG×E studies were significant compared with 27% of replication attempts. These findings are consistent with the existence of publication bias among novel cG×E studies, making cG×E hypotheses appear more robust than they actually are. There also appears to be publication bias among replication attempts because positive replication attempts had smaller average sample sizes than negative ones. Power calculations using observed sample sizes suggest that cG×E studies are underpowered. Low power along with the likely low prior probability of a given cG×E hypothesis being true suggests that most or even all positive cG×E findings represent type I errors.In this new era of big data and small effects, a recalibration of views about groundbreaking findings is necessary. Well-powered direct replications deserve more attention than novel cG×E findings and indirect replications.
Research review: Gene- environment interaction research in youth depression-A systematic review with recommendations for future research
Biological sensitivity to context
The role of stressful parenting and mineralocorticoid receptor haplotypes on social development during adolescence and young adulthood
DOI:10.1007/s10964-019-00988-2
PMID:30805852
[本文引用: 1]
The development of social behavior could be affected by stressful parenting. The mineralocorticoid receptor, one of the two main receptors for the stress hormone cortisol, plays a vital role in adequate responses to stress. Therefore, the effects of stressful parenting on social development (i.e., empathic concern, perspective taking and prosocial behavior) may be moderated by functional genetic variation in mineralocorticoid receptor haplotypes (a combination of alleles). A group of 343 adolescents (44.3% females) was followed from the age of 13 until 24 years. Growth curve analyses showed lower levels of prosocial behaviors and a slower increase in empathic concern and perspective taking in adolescents who reported more stressful parenting. In contrast, relatively higher levels of prosocial behavior, empathic concern and perspective taking were present in combination with stress resilient mineralocorticoid receptor haplotypes. Despite sex differences in social development with earlier social development for girls, no consistent sex differences were found with regard to mineralocorticoid receptor haplotypes. The current study showed that genetic variation in mineralocorticoid receptor impacts the social development during adolescence and young adulthood.
Preschool children’s prosocial behavior: The role of mother-child, father-child and teacher-child relationships
HPA axis genes, and their interaction with childhood maltreatment, are related to cortisol levels and stress- related phenotypes
DOI:10.1038/npp.2017.118
PMID:28589964
[本文引用: 1]
Stress responses are controlled by the hypothalamus pituitary adrenal (HPA)-axis and maladaptive stress responses are associated with the onset and maintenance of stress-related disorders such as major depressive disorder (MDD). Genes that play a role in the HPA-axis regulation may likely contribute to the relation between relevant neurobiological substrates and stress-related disorders. Therefore, we performed gene-wide analyses for 30 a priori literature-based genes involved in HPA-axis regulation in 2014 subjects (34% male; mean age: 42.5) to study the relations with lifetime MDD diagnosis, cortisol awakening response, and dexamethasone suppression test (DST) levels (subsample N=1472) and hippocampal and amygdala volume (3T MR images; subsample N=225). Additionally, gene by childhood maltreatment (CM) interactions were investigated. Gene-wide significant results were found for dexamethasone suppression (CYP11A1, CYP17A1, POU1F1, AKR1D1), hippocampal volume (CYP17A1, CYP11A1, HSD3B2, PROP1, AVPRA1, SRD5A1), amygdala volume (POMC, CRH, HSD3B2), and lifetime MDD diagnosis (FKBP5 and CRH), all permutation p-values<0.05. Interactions with CM were found for several genes; the strongest interactions were found for NR3C2, where the minor allele of SNP rs17581262 was related to smaller hippocampal volume, smaller amygdala volume, higher DST levels, and higher odds of MDD diagnosis only in participants with CM. As hypothesized, several HPA-axis genes are associated with stress-related endophenotypes including cortisol response and reduced brain volumes. Furthermore, we found a pleiotropic interaction between CM and the mineralocorticoid receptor gene, suggesting that this gene plays an important moderating role in stress and stress-related disorders.
Moderation of adult depression by the serotonin transporter promoter variant (5-HTTLPR), childhood abuse and adult traumatic events in a general population sample
Measurement of deliberate self-harm: Preliminary data on the Deliberate Self-Harm Inventory
Cultural conceptions of learning and development
In
Behavioral genetic analyses of prosocial behavior in adolescents
DOI:10.1111/j.1467-7687.2008.00739.x
PMID:19120424
[本文引用: 1]
Prosocial behavior is an important aspect of normal social and psychological development. Adult and child twin studies typically estimate the heritability of prosocial behavior to be between 30 and 50%, although relatively little is known about genetic and environmental influences upon prosocial behavior in adolescence. We therefore examined reports of prosocial behavior in a large longitudinal family study of 1160 adolescent twin pairs (aged between 13 and 19 years). Prosocial behavior was assessed at two time points by self-report and at the second time point by additional parent-ratings using the Strengths and Difficulties Questionnaire (SDQ; Goodman, 1997). Adolescent females were reported to be significantly more prosocial than males (p <.001). Univariate analyses primarily showed moderate heritability and large nonshared environmental influences. There was a moderate genetic correlation between self- and parent-reported prosocial behaviour, suggesting that both types of rater were tapping into genetically overlapping constructs. Longitudinal analyses revealed that continuity was largely explained by genes. Unique environmental influences were predominantly time-specific and were the major source of individual differences.
The neurobiology of stress and development
Stress is a part of every life to varying degrees, but individuals differ in their stress vulnerability. Stress is usefully viewed from a biological perspective; accordingly, it involves activation of neurobiological systems that preserve viability through change or allostasis. Although they are necessary for survival, frequent neurobiological stress responses increase the risk of physical and mental health problems, perhaps particularly when experienced during periods of rapid brain development. Recently, advances in noninvasive measurement techniques have resulted in a burgeoning of human developmental stress research. Here we review the anatomy and physiology of stress responding, discuss the relevant animal literature, and briefly outline what is currently known about the psychobiology of stress in human development, the critical role of social regulation of stress neurobiology, and the importance of individual differences as a lens through which to approach questions about stress experiences during development and child outcomes.
Computational procedures for probing interactions in OLS and logistic regression: SPSS and SAS implementations
DOI:10.3758/BRM.41.3.924
PMID:19587209
[本文引用: 1]
Researchers often hypothesize moderated effects, in which the effect of an independent variable on an outcome variable depends on the value of a moderator variable. Such an effect reveals itself statistically as an interaction between the independent and moderator variables in a model of the outcome variable. When an interaction is found, it is important to probe the interaction, for theories and hypotheses often predict not just interaction but a specific pattern of effects of the focal independent variable as a function of the moderator. This article describes the familiar pick-a-point approach and the much less familiar Johnson-Neyman technique for probing interactions in linear models and introduces macros for SPSS and SAS to simplify the computations and facilitate the probing of interactions in ordinary least squares and logistic regression. A script version of the SPSS macro is also available for users who prefer a point-and-click user interface rather than command syntax.
The link between childhood trauma and depression: Insights from HPA axis studies in humans
DOI:10.1016/j.psyneuen.2008.03.008
PMID:18602762
[本文引用: 1]
Childhood trauma is a potent risk factor for developing depression in adulthood, particularly in response to additional stress. We here summarize results from a series of clinical studies suggesting that childhood trauma in humans is associated with sensitization of the neuroendocrine stress response, glucocorticoid resistance, increased central corticotropin-releasing factor (CRF) activity, immune activation, and reduced hippocampal volume, closely paralleling several of the neuroendocrine features of depression. Neuroendocrine changes secondary to early-life stress likely reflect risk to develop depression in response to stress, potentially due to failure of a connected neural circuitry implicated in emotional, neuroendocrine and autonomic control to compensate in response to challenge. However, not all of depression is related to childhood trauma and our results suggest the existence of biologically distinguishable subtypes of depression as a function of childhood trauma that are also responsive to differential treatment. Other risk factors, such as female gender and genetic dispositions, interfere with components of the stress response and further increase vulnerability for depression. Similar associations apply to a spectrum of other psychiatric and medical disorders that frequently coincide with depression and are aggravated by stress. Taken together, this line of evidence demonstrates that psychoneuroendocrine research may ultimately promote optimized clinical care and help prevent the adverse outcomes of childhood trauma.
Linking individual variability in functional brain connectivity to polygenic risk in major depressive disorder
The effects of the parent- child relationship and parental educational involvement on adolescent depression, self-injury, and suicidal ideation: The roles of defeat and meaning in life
亲子关系和父母教育卷入对青少年抑郁、自伤和自杀意念的影响: 挫败感和人生意义感的作用
DOI:10.3724/SP.J.1041.2023.00129
[本文引用: 2]
基于生态系统理论和动机?意志整合模型, 采用问卷法以930名青少年(年龄 = 15.24 ± 1.66岁)及其父母为研究对象, 构建一个有调节的中介模型, 从家长与孩子双视角分析孩子体验到的亲子关系和父母教育卷入对青少年抑郁、自伤和自杀意念的影响路径。结果显示:(1)相比低亲子关系?低教育卷入一致的个体, 高亲子关系?高教育卷入的青少年有着更低水平的挫败感, 且相比于低亲子关系?高教育卷入的青少年, 有着高亲子关系?低教育卷入的个体表现出更低水平的挫败感; (2)挫败感在亲子关系?教育卷入与青少年抑郁、自伤和自杀意念之间起部分中介作用; (3)人生意义感调节该中介模型后半段, 即挫败感对青少年抑郁、自伤和自杀意念的影响, 具体表现为随着人生意义感的增加, 挫败感对抑郁、自伤和自杀意念的影响逐渐减小。研究从家庭关系中家长与孩子双视角, 揭示了挫败感和人生意义感的中介与调节作用, 为青少年抑郁、自伤和自杀意念的发生机制提供更多解释路径。
Childhood quality influences genetic sensitivity to environmental influences across adulthood: A life-course gene × environment interaction study
DOI:10.1017/S0954579417001493
PMID:29162193
[本文引用: 4]
While environmental adversity has been shown to increase risk for psychopathology, individuals differ in their sensitivity to these effects. Both genes and childhood experiences are thought to influence sensitivity to the environment, and these factors may operate synergistically such that the effects of childhood experiences on later sensitivity are greater in individuals who are more genetically sensitive. In line with this hypothesis, several recent studies have reported a significant three-way interaction (Gene × Environment × Environment) between two candidate genes and childhood and adult environment on adult psychopathology. We aimed to replicate and extend these findings in a large, prospective multiwave longitudinal study using a polygenic score of environmental sensitivity and objectively measured childhood and adult material environmental quality. We found evidence for both Environment × Environment and Gene × Environment × Environment effects on psychological distress. Children with a poor-quality material environment were more sensitive to the negative effects of a poor environment as adults, reporting significantly higher psychological distress scores. These effects were further moderated by a polygenic score of environmental sensitivity. Genetically sensitive children were more vulnerable to adversity as adults, if they had experienced a poor childhood environment but were significantly less vulnerable if their childhood environment was positive. These findings are in line with the differential susceptibility hypothesis and suggest that a life course approach is necessary to elucidate the role of Gene × Environment in the development of mental illnesses.
Gene× environment interaction studies have not properly controlled for potential confounders: The problem and the (simple) solution
Child temperament moderates effects of parent-child mutuality on self‐regulation: A relationship-based path for emotionally negative infants
Revisioning Prosocial Tendencies Measure for Adolescent
青少年亲社会倾向量表的修订
The differential relation of childhood maltreatment to stress sensitization in adolescent and young adult depression
DOI:10.1016/j.adolescence.2014.05.012
PMID:24960433
[本文引用: 1]
Childhood maltreatment has been shown to have a stronger etiological relation to depression onset in adolescence than in adulthood. We propose that a maltreatment history may more strongly sensitize individuals to the depressogenic effects of proximal stressful life events in adolescence compared to adulthood. In an amalgamated sample of 176 unipolar depressed adolescents (age 12-17) and emerging adults (age 18-29), we examined the moderating role of age group on the relation of childhood maltreatment to sensitization to stressors that occurred just prior to episode onset. Among adolescents, but not among adults, those with a maltreatment history reported a lower severity level of life events prior to episode onset than reported by those without such a history. Further, this relation was specific to emotional abuse, and not physical or sexual abuse. We suggest that the pathological mechanisms associated with translating childhood maltreatment to depression may differ across developmental periods. Copyright © 2014 The Foundation for Professionals in Services for Adolescents. Published by Elsevier Ltd. All rights reserved.
Parent-child relationship quality and emotional adjustment among college students: The role of parental financial support
Mother-child and father-child mutuality in two contexts: Consequences for young children's peer relationships
Childhood maltreatment and non-suicidal self-injury: A systematic review and meta-analysis
Representational and questionnaire measures of attachment: A meta-analysis of relations to child internalizing and externalizing problems
DOI:10.1037/bul0000029
PMID:26619212
[本文引用: 1]
Although the quality of the attachment relationship is often cited as an important determinant of development, the extent of impact of this environmental influence in shaping behavioral outcomes has been a matter of considerable debate. This may, in part, be because of the variability in methodologies used for assessing attachment across infancy, childhood, and adolescence, including behavioral, representational, and questionnaire measures of attachment. Previous meta-analyses of the relations between attachment and internalizing and externalizing problems have focused on the behavioral measures of attachment used primarily in infancy. The current meta-analysis is a comprehensive examination of the literature on attachment and behavioral problems in children aged 3-18 years, focusing on the representational and questionnaire measures most commonly used in this age range. When secure attachment was compared with insecure attachment, modest associations with internalizing behavior (165 studies; 48,224 families; d =.58; 95% confidence interval [CI] [.52-.64]) were found. Multivariate moderator analyses were used to disentangle the unique influence of each significant univariate moderator more precisely, and results revealed that effect sizes decreased as the child aged, and were larger in studies in which the participants were ethnically White, where the child was the problem informant, and when the internalizing measure was depressive symptoms. Attachment and externalizing behavior were also associated (116 studies; 24,689 families; d =.49; 95% CI [42-.56]), and effect sizes were larger in ethnically White samples, and in those where the child was the problem informant. Avoidant, ambivalent, and disorganized attachment classifications were associated with internalizing behavior, but only disorganized attachment was associated with externalizing behavior.(c) 2016 APA, all rights reserved).
The neurobiology of oppositional defiant disorder and conduct disorder: Altered functioning in three mental domains
DOI:10.1017/S0954579412000272
PMID:22800761
[本文引用: 1]
This review discusses neurobiological studies of oppositional defiant disorder and conduct disorder within the conceptual framework of three interrelated mental domains: punishment processing, reward processing, and cognitive control. First, impaired fear conditioning, reduced cortisol reactivity to stress, amygdala hyporeactivity to negative stimuli, and altered serotonin and noradrenaline neurotransmission suggest low punishment sensitivity, which may compromise the ability of children and adolescents to make associations between inappropriate behaviors and forthcoming punishments. Second, sympathetic nervous system hyporeactivity to incentives, low basal heart rate associated with sensation seeking, orbitofrontal cortex hyporeactiviy to reward, and altered dopamine functioning suggest a hyposensitivity to reward. The associated unpleasant emotional state may make children and adolescents prone to sensation-seeking behavior such as rule breaking, delinquency, and substance abuse. Third, impairments in executive functions, especially when motivational factors are involved, as well as structural deficits and impaired functioning of the paralimbic system encompassing the orbitofrontal and cingulate cortex, suggest impaired cognitive control over emotional behavior. In the discussion we argue that more insight into the neurobiology of oppositional defiance disorder and conduct disorder may be obtained by studying these disorders separately and by paying attention to the heterogeneity of symptoms within each disorder.
Childhood adversity, adult stressful life events, and risk of past-year psychiatric disorder: A test of the stress sensitization hypothesis in a population-based sample of adults
DOI:10.1017/S0033291709992121
PMID:20018126
[本文引用: 1]
Childhood adversity (CA) is associated with adult mental disorders, but the mechanisms underlying this association remain inadequately understood. Stress sensitization, whereby CA increases vulnerability to mental disorders following adult stressful life events, has been proposed as a potential mechanism. We provide a test of the stress sensitization hypothesis in a national sample.We investigated whether the association between past-year stressful life events and the 12-month prevalence of major depression, post-traumatic stress disorder (PTSD), other anxiety disorders, and perceived stress varies according to exposure to CA. We used data from the National Epidemiological Survey of Alcohol and Related Conditions (NESARC) (n=34 653).Past-year stressful life events were associated with an increased risk of major depression, PTSD, anxiety disorders, and perceived stress. However, the magnitude of the increased risk varied according to respondents' history of CA. For example, past-year major stressors were associated with a 27.3% increase in the 12-month risk of depression among individuals with 3 CAs and a 14.8% increased risk among individuals without CAs. Stress sensitization effects were present for depression, PTSD, and other anxiety disorders in women and men, although gender differences were found in the threshold of past-year stress needed to trigger such effects. Stress sensitization was most evident among individuals with 3 CAs.CA is associated with increased vulnerability to the deleterious mental health effects of adult stressors in both men and women. High levels of CA may represent a general diathesis for multiple types of psychopathology that persists throughout the life course.
DNA methylation, early life environment, and health outcomes
Trauma, helpfulness and selfishness: The effect of abuse and neglect on altruistic, moral and pro-social capacities
The colour naming of socially threatening words
Stress-system genes and life stress predict cortisol levels and amygdala and hippocampal volumes in children
DOI:10.1038/npp.2013.327
PMID:24304824
[本文引用: 2]
Depression has been linked to increased cortisol reactivity and differences in limbic brain volumes, yet the mechanisms underlying these alterations are unclear. One main hypothesis is that stress causes these effects. This is supported by animal studies showing that chronic stress or glucocorticoid administration can lead to alterations in hippocampal and amygdala structures. Relatedly, life stress is cited as one of the major risk factors for depression and candidate gene studies have related variation in stress-system genes to increased prevalence and severity of depression. The present study tested the hypothesis that genetic profile scores combining variance across 10 single nucleotide polymorphisms from four stress-system genes (CRHR1, NR3C2, NR3C1, and FKBP5) and early life stress would predict increases in cortisol levels during laboratory stressors in 120 preschool-age children (3-5 years old), as well as hippocampal and amygdala volumes assessed with MRI in these same children at school age (7-12 years old). We found that stress-system genetic profile scores positively predicted cortisol levels while the number of stressful/traumatic life events experienced by 3-5 years old negatively predicted cortisol levels. The interaction of genetic profile scores and early life stress predicted left hippocampal and left amygdala volumes. Cortisol partially mediated the effects of genetic variation and life stress on limbic brain volumes, particularly on left amygdala volume. These results suggest that stress-related genetic and early environmental factors contribute to variation in stress cortisol reactivity and limbic brain volumes in children, phenotypes associated with depression in adulthood.
Dictator game giving: The importance of descriptive versus injunctive norms
Childhood emotional abuse, physical abuse, and neglect are associated with theory of mind decoding accuracy in young adults with depression
DOI:S0165-1781(17)32232-1
PMID:30165325
[本文引用: 1]
The current study is the first to examine the relation of childhood abuse and neglect history to theory of mind decoding accuracy as moderated by depression. Fifty-five young adults with current or lifetime unipolar depression diagnosis and 70 never-depressed young adults completed the 'Reading the Mind in the Eyes task,' (RMET). Childhood emotional abuse, physical abuse, and neglect were assessed with a gold-standard contextual interview with standardized, independent ratings. Poorer RMET accuracy was associated with a history of emotional abuse in the depressed group and a history of physical abuse in the non-depressed group. In contrast, across both groups, those with a history of neglect showed significantly enhanced theory of mind decoding accuracy compared to those without. Further, differential accuracy across positive, negative, and neutral valenced stimuli in the RMET was observed in each model. These findings indicate that distinct theory of mind performance results from early experiences of threat versus deprivation, and suggest that early intervention may be most successful in preventing negative interpersonal outcomes of maltreatment by focusing on remediating theory of mind deficits resulting from abuse, and tempering heightened sensitivity in those exposed to neglect.Copyright © 2018 Elsevier B.V. All rights reserved.
Distinguishing differential susceptibility from diathesis-stress: Recommendations for evaluating interaction effects
DOI:10.1017/S0954579412000065
PMID:22559121
[本文引用: 1]
This report describes the state of the art in distinguishing data generated by differential susceptibility from diathesis-stress models. We discuss several limitations of existing practices for probing interaction effects and offer solutions that are designed to better differentiate differential susceptibility from diathesis-stress models and quantify their corresponding implications. In addition, we demonstrate the utility of these methods by revisiting published evidence suggesting that temperamental difficulty serves as a marker of enhanced susceptibility to early maternal caregiving across a range of outcome domains in the NICHD Study of Early Child Care and Youth Development. We find that, with the exception of mother reports of psychopathology, there is consistent evidence in the Study of Early Child Care and Youth Development that the predictive significance of early sensitivity is moderated by difficult temperament over time. However, differential susceptibility effects emerged primarily for teacher reports of academic skills, social competence, and symptomatology. In contrast, effects more consistent with the diathesis-stress model were obtained for mother reports of social skills and objective tests of academic skills. We conclude by discussing the value of the application of this work to the next wave of Gene × Environment studies focused on early caregiving experiences.
MR and GR functional SNPs may modulate tobacco smoking susceptibility
DOI:10.1007/s00702-013-1012-2
PMID:23543128
[本文引用: 2]
A number of studies have demonstrated that stress is involved in all aspects of smoking behavior, including initiation, maintenance and relapse. The mineralocorticoid (MR) and glucocorticoid (GR) receptors are expressed in several brain areas and play a key role in negative feedback of the hypothalamic-pituitary-adrenal (HPA) axis. As nicotine increases the activation of the HPA axis, we wondered if functional SNPs (single nucleotide polymorphisms) in MR and GR coding genes (NR3C2 rs5522 and NR3C1 rs6198, respectively) may be involved in smoking susceptibility. The sample included 627 volunteers, of which 514 were never-smokers and 113 lifetime smokers. We report an interaction effect between rs5522 and rs6198 SNPs. The odds ratio (OR) for the presence of the NR3C2 rs5522 Val allele in NR3C1 rs6198 G carriers was 0.18 (P = 0.007), while in rs6198 G noncarriers the OR was 1.83 (P = 0.027). We also found main effects of the NR3C1 rs6198 G allele on number of cigarettes smoked per day (P = 0.027) and in total score of the Fagerström Test for Nicotine Dependence (P = 0.007). These findings are consistent with a possible link between NR3C2 and NR3C1 polymorphisms and smoking behavior and provide a first partial replication for a nominally significant GWAS finding between NR3C2 and tobacco smoking.
The role of childhood abuse for suicidality in the context of the interpersonal theory of suicide: An investigation in German psychiatric inpatients with depression
DOI:S0165-0327(18)31522-2
PMID:30448764
[本文引用: 1]
Several studies provide evidence for a relationship between childhood abuse and suicidality across the lifespan. To examine this association in the context of the Interpersonal Psychological Theory of Suicide (IPTS), we investigated whether its constructs thwarted belongingness, perceived burdensomeness and capability for suicide are potential mediators.Eighty-four German psychiatric inpatients with unipolar depression (M = 37.6 years, 69% female) and current or lifetime suicidal ideation were included. For the assessment we used the Childhood Trauma Screener (CTS), the Rasch-based Screening for Depression (DESC-I), the Interpersonal Needs Questionnaire (INQ), the German Capability for Suicide Questionnaire (GCSQ), the Beck Scale for Suicide Ideation (BSS) and the Suicide Behaviors Questionnaire-Revised (SBQ-R). Simple and multiple mediator analyses were applied.Most patients (70%) had experienced childhood abuse. Emotional abuse showed an indirect association with suicidal ideation via thwarted belongingness and perceived burdensomeness, whereas physical and sexual abuse were indirectly related to suicide risk via capability for suicide.The small sample size and the cross-sectional design are limiting factors of the present study.Childhood abuse is a common experience of inpatients with unipolar depression. This study showed its indirect effects on suicidal ideation and risk for suicide, mediated by the constructs of the IPTS. Further research should investigate this issue in other populations and clinicians should be aware of the devastating effects of childhood abuse.Copyright © 2018. Published by Elsevier B.V.
Diurnal cortisol interacts with stressful events to prospectively predict depressive symptoms in adolescent girls
Genetic susceptibility to parenting style: DRD2 and COMT influence creativity
Sensitizing effect of early adversity on depressive reactions to later proximal stress: Moderation by polymorphisms in serotonin transporter and corticotropin releasing hormone receptor genes in a 20-year longitudinal study
HPA-axis multilocus genetic variation moderates associations between environmental stress and depressive symptoms among adolescents
DOI:10.1017/S0954579418000779
PMID:30394263
[本文引用: 8]
Research suggests that genetic variants linked to hypothalamic-pituitary-adrenal (HPA)-axis functioning moderate the association between environmental stressors and depression, but examining gene-environment interactions with single polymorphisms limits power. The current study used a multilocus genetic profile score (MGPS) approach to measuring HPA-axis-related genetic variation and examined interactions with acute stress, chronic stress, and childhood adversity (assessed using contextual threat interview methods) with depressive symptoms as outcomes in an adolescent sample (ages 14-17, N = 241; White subsample n = 192). Additive MGPSs were calculated using 10 single nucleotide polymorphisms within HPA-axis genes (CRHR1, NR3C2, NR3C1, FKBP5). Higher MGPS directly correlated with adolescent depressive symptoms. Moreover, MGPS predicted stronger associations between acute and chronic stress and adolescent depressive symptoms and also moderated the effect of interpersonal, but not noninterpersonal, childhood adversity. Gene-environment interactions individually accounted for 5%-8% of depressive symptom variation. All results were retained following multiple test correction and stratification by race. Results suggest that using MGPSs provides substantial power to examine gene-environmental interactions linked to affective outcomes among adolescents.
Stress sensitization to depression following childhood adversity: Moderation by HPA axis and serotonergic multilocus profile scores
Individual differences in early adolescents’ latent trait cortisol (LTC): Relation to early adversity
The early-life social environment and DNA methylation
DOI:10.1111/j.1399-0004.2012.01843.x
PMID:22236068
[本文引用: 1]
DNA methylation is a chemical modification of DNA that confers, upon identical sequences, different identities that are reflected in different gene expression programming. DNA methylation has a well-established role in cellular differentiation by providing a mechanism for one genome to express multiple phenotypes in a multicellular organism. Recent data point however to the possibility that in addition to the innate process of cellular differentiation, DNA methylation can serve as a genome adaptation mechanism, adapting genome function to changing environmental contexts including social environments. A critical time point for this process is early life when cues from the social and physical environments define lifelong trajectories of physical and mental health. DNA methylation and additional epigenetic modifications could therefore serve as molecular links between 'nurture' and 'nature'. Data that are consistent with this new role for DNA methylation as a mechanism for conferring an 'environment' specific identity to DNA will be discussed.© 2012 John Wiley & Sons A/S.
The role of adverse childhood experience on depression symptom, prevalence, and severity among school going adolescents
Child maltreatment and sleep problems among adolescents in Ontario: A cross sectional study
The rs7208505 polymorphism and differential expression of the SKA 2 gene in the prefrontal cortex of suicide victims from the Mexican population
DOI:10.1080/13811118.2023.2209155
URL
PMID:37204142
[本文引用: 1]
The main aim of the current study was to investigate whether 2 gene expression in the postmortem brain of rs7208505 genotype are altered in suicide victims from a Mexican population.In this study, we report a genetic analysis of expression levels of the 2 gene in the prefrontal cortex of the postmortem brain of suicidal subjects ( = 22) compared to subjects who died of causes other than suicide ( = 22) in a Mexican population using RT-qPCR assays. Additionally, we genotyped the rs7208505 polymorphism in suicide victims ( = 98) and controls ( = 88) and we evaluate the association of genotypes for the SNP rs7208505 with expression level of 2.The results showed that the expression of the 2 gene was significantly higher in suicide victims compared to control subjects ( = 0.044). Interestingly, we observed a greater proportion of allele A of the rs7208505 in suicide victims than controls. Even though there was no association between the SNP with suicide in the study population we found a significative association of the expression level from 2 with the allele A of the rs7208505 and suicide.The evidence suggests that the expression of 2 in the prefrontal cortex may be a critical factor in the etiology of suicidal behavior.
A meta-analysis on interparental conflict, parenting, and child adjustment in divorced families: Examining mediation using meta-analytic structural equation models
Mineralocorticoid receptor haplotypes sex-dependently moderate depression susceptibility following childhood maltreatment
DOI:10.1016/j.psyneuen.2015.01.018
PMID:25686805
[本文引用: 2]
The MR is an important regulator of the hypothalamic-pituitary-adrenal (HPA) axis and a prime target for corticosteroids. There is increasing evidence from both clinical and preclinical studies that the MR has different effects on behavior and mood in males and females. To investigate the hypothesis that the MR sex-dependently influences the relation between childhood maltreatment and depression, we investigated three common and functional MR haplotypes (GA, CA, and CG haplotype, based on rs5522 and rs2070951) in a population-based cohort (N = 665) and an independent clinical cohort from the Netherlands Study of Depression and Anxiety (NESDA) (N = 1639). The CA haplotype sex-dependently moderated the relation between childhood maltreatment and depressive symptoms both in the population-based sample (sex × maltreatment × haplotype: β = -4.07, P = 0.029) and in the clinical sample (sex × maltreatment × haplotype, β = -2.40, P = 0.011). Specifically, female individuals in the population-based sample were protected (β = -4.58, P = 2.0 e(-5)), whereas males in the clinical sample were at increased risk (β = 2.54, P = 0.0022). In line with these results, female GA haplotype carriers displayed increased vulnerability in the population-based sample (β = 4.58, P = 7.5 e(-5)) whereas male CG-carriers showed increased resilience in the clinical sample (β = -2.71, P = 0.016). Consistently, we found a decreased lifetime MDD risk for male GA haplotype carriers following childhood maltreatment but an increased risk for male CA haplotype carriers in the clinical sample. In both samples, sex-dependent effects were observed for GA-GA diplotype carriers. In summary, sex plays an important role in determining whether functional genetic variation in MR is beneficial or detrimental, with an apparent female advantage for the CA haplotype but male advantage for the GA and CG haplotype. These sex-dependent effects of MR on depression susceptibility following childhood maltreatment are relevant in light of the increased prevalence of mood disorders in women and point to a sex-specific role of MR in the etiology of depression following childhood maltreatment.Copyright © 2015 Elsevier Ltd. All rights reserved.
Stress sensitization to depression following childhood adversity: Moderation by HPA axis and serotonergic multilocus profile scores
Additive genetic risk from five serotonin system polymorphisms interacts with interpersonal stress to predict depression
DOI:10.1037/abn0000098
PMID:26595467
[本文引用: 1]
Behavioral genetic research supports polygenic models of depression in which many genetic variations each contribute a small amount of risk, and prevailing diathesis-stress models suggest gene-environment interactions (G×E). Multilocus profile scores of additive risk offer an approach that is consistent with polygenic models of depression risk. In a first demonstration of this approach in a G×E predicting depression, we created an additive multilocus profile score from 5 serotonin system polymorphisms (1 each in the genes HTR1A, HTR2A, HTR2C, and 2 in TPH2). Analyses focused on 2 forms of interpersonal stress as environmental risk factors. Using 5 years of longitudinal diagnostic and life stress interviews from 387 emerging young adults in the Youth Emotion Project, survival analyses show that this multilocus profile score interacts with major interpersonal stressful life events to predict major depressive episode onsets (hazard ratio [HR] = 1.815, p =.007). Simultaneously, there was a significant protective effect of the profile score without a recent event (HR = 0.83, p =.030). The G×E effect with interpersonal chronic stress was not significant (HR = 1.15, p =.165). Finally, effect sizes for genetic factors examined ignoring stress suggested such an approach could lead to overlooking or misinterpreting genetic effects. Both the G×E effect and the protective simple main effect were replicated in a sample of early adolescent girls (N = 105). We discuss potential benefits of the multilocus genetic profile score approach and caveats for future research.(c) 2015 APA, all rights reserved).
Theory Models on Gene-environment Interaction
DOI:10.3724/SP.J.1042.2015.01852
[本文引用: 1]
There are three representative theory models about gene-environment interaction: diathesis-stress model, differential susceptibility model and vantage sensitivity model. Diathesis-stress model advocates that one who stays near ink gets stained black, vantage sensitivity model, however, suggests that one who stays near vermilion gets stained red, and differential susceptibility model is eclectic. To verify the above-mentioned model has been one of the focuses of research on gene-environment interaction. The grouping regression analysis and hierarchical regression analysis are the traditional methods to test these three models, and the region of significance and the re-parameterized regression model are the newly emerging ones. Future research could focus on the issues of domain-specificity, the ethnic difference, developmental change and the underlying mechanism of the gene-environment interaction. The measurement and method to examine the interaction between gene and environment also remain improved.
基因-环境交互作用理论模型及其检验方法
DOI:10.3724/SP.J.1042.2015.01852
[本文引用: 1]
素质?压力模型、不同易感性模型和优势敏感性模型是当前有关基因?环境交互作用的三种代表性理论模型。素质?压力模型认为“近墨者黑”或“出淤泥而不染”, 优势反应敏感性模型认为“近朱者赤”, 而不同易感性模型则兼收并蓄, 认为某些个体近墨则黑, 近朱则赤。检验上述模型的现实有效性是当前基因?环境交互作用研究领域的热点问题之一。概观而言, 分组回归和分层回归是常用的传统检验方法, 显著性区域分析法和新参数回归模型法则是新近兴起的。未来研究需要进一步探索三种模型的领域特殊性、种族差异等问题, 检验这些模型的方法也有待改善。
Association between negative life events and early adolescents’ depression: The moderating effects of Catechol-O-methyltransferase (COMT) Gene Val158Met polymorphism and parenting behavior
负性生活事件与青少年早期抑郁的关系:COMT基因Val158Met多态性与父母教养行为的调节作用
DOI:10.3724/SP.J.1041.2019.00903
[本文引用: 1]
采用环境×基因×环境(E×G×E)研究设计, 以637名青少年为被试, 考察了负性生活事件、COMT基因Val158Met多态性和父母教养行为对青少年早期抑郁的影响。结果发现:负性生活事件对青少年早期抑郁具有显著正向预测作用, 且COMT基因Val158Met多态性和父亲积极教养行为在其中起调节作用, 但该调节作用仅存在于男青少年群体中:在携带Val/Val基因型的男青少年中, 当父亲积极教养行为水平较低时, 青少年的抑郁水平随负性生活事件的增多而显著上升, 当父亲积极教养行为水平较高时, 负性生活事件对抑郁无显著预测作用; 在携带Met等位基因的男青少年中, 上述交互作用不显著。
Relations between shyness and psychological adjustment in Chinese children: The role of friendship quality
The relationship between personality types and prosocial behavior and aggression in Chinese adolescents
Factor structure and criterion validity across the full scale and ten short forms of the CES-D among Chinese adolescents
DOI:10.1037/pas0000559
PMID:29658726
[本文引用: 1]
We systematically examined the factor structure and criterion validity across the full scale and 10 short forms of the Center for Epidemiological Studies Depression Scale (CES-D) with Chinese youth. Participants were 5,434 Chinese adolescents in Grades 7 to 12 who completed the full CES-D; 612 of them further completed a structured diagnostic interview with the major depressive disorder (MDD) module of the Kiddie Schedule for Affective Disorder and Schizophrenia for School-age Children. Using a split-sample approach, a series of 4-, 3-, 2-, and 1-factor models were tested using exploratory structural equation modeling and cross-validated using confirmatory factor analysis; the dimensionality was also evaluated by parallel analysis in conjunction with the scree test and aided by factor mixture analysis. The results indicated that a single-factor model of depression with a wording method factor fitted the data well, and was the optimal structure underlying the scores of the full and shortened CES-D. Additionally, receiver operating characteristic curve analyses for MDD case detection showed that the CES-D full-scale scores accurately detected MDD youth (area under the curve [AUC] =.84). Furthermore, the short-form scores produced comparable AUCs with the full scale (.82 to.85), as well as similar levels of sensitivity and specificity when using optimal cutoffs. These findings suggest that depression among Chinese adolescents can be adequately measured and screened for by a single-factor structure underlying the CES-D scores, and that the short forms provide a viable alternative to the full instrument. (PsycINFO Database Record(c) 2018 APA, all rights reserved).
书面调查和网络调查的区别——两种数据收集方法的比较
Three phases of Gene × Environment interaction research: Theoretical assumptions underlying gene selection
The effects of NR3C1 polymorphisms and paternal/maternal parenting styles on Chinese adolescent anxiety disorders
NR3C1基因多态性及单倍型、父母教养方式对青少年焦虑障碍的影响
Interaction of FKBP5 gene variants and adverse life events in predicting depression onset: Results from a 10-year prospective community study
DOI:10.1176/appi.ajp.2011.10111577
PMID:21865530
[本文引用: 1]
The binding protein FKBP5 is an important modulator of the function of the glucocorticoid receptor, the main receptor of the stress hormone system. This turns the FKBP5 gene into a key candidate for gene-environment interactions, which are considered critical for pathogenesis of stress-related disorders. The authors explored gene-environment interactions between FKBP5 gene variants and adverse life events in predicting the first occurrence of a major depressive episode.The analyses were based on 884 Caucasians in a 10-year prospective community study. At baseline, they were 14-24 years old and did not fulfill criteria for a major depressive episode. The DSM-IV-based Munich Composite International Diagnostic Interview was used to assess adverse life events preceding baseline and major depressive episodes during follow-up. On the basis of previous findings, five single-nucleotide polymorphisms (SNPs) within the FKBP5 gene were selected for genotyping.While the authors did not observe genetic main effects, they found interactions between the five SNPs and traumatic (but not separation) events, with the strongest effect for severe trauma. The effect of trauma on incident major depressive episodes was evident among subjects homozygous for the minor alleles but not subjects with other genotypes. The findings were replicated in the U.K. Environmental Risk Longitudinal Twin Study.These hypothesis-driven results suggest that an interaction between FKBP5 genotype and trauma is involved in the onset of depression. Subjects homozygous for the minor alleles of the investigated FKBP5 SNPs seem to be particularly sensitive to effects of trauma exposure in terms of triggering depression onset.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}