

1 前言
以往研究者针对动作指向物理客体以满足其需求的非社会性意图(即客体导向性意图, object- directed intention, 其不涉及他人)的识别, 开展了大量研究(Heineman-Pieper, 2009), 提出视觉系统将行为场景分为动作、目标(或结果)状态和情境因素三个基本要素; 个体对这三个要素间关系的解释, 遵循动作在情境因素的约束下以最小成本实现目标状态这一最根本的理性原则(Csibra et al., 2003; Gergely & Csibra, 2003)。Jara-Ettinger等采用效用函数(Utility = Reward - Cost)来描述动作与目标的关系, 其认为由于行为主体具备理性这一根本特性, 故动作遵循效用最大化的方式实现目标。因此, 当其它条件不变时, 若行为和当前所趋近的对象具有最大效用值时, 则两者间具有目标指向关系(Jara-Ettinger et al., 2015)。然而, 动作意图不仅包括以指向物理客体为目标的客体导向性意图, 还包含以指向社会主体为目标, 且影响对方行为的社会交互意图(Canessa et al., 2012; Hobbs & Spelke, 2015), 也即社会性意图(social intention)。针对社会性意图的识别(主要是指两人的交互, 下同), 有研究者提出双方行为的耦合性(contingency)是关键决定因素之一, 认为只要某一个体动作导致另一个体动作改变且具有即时同步性, 二者间就可能被视为存在以交互为目的的指向关系(详见de Jaegher et al., 2010的综述), 如:握手。然而, 后续研究表明, 当两个个体共同追逐的某个目标被设置为不可见时, 即使个体间行为存在耦合性, 人们也难以识别二者间的意图; 只有当追逐的目标可见时, 其行为才被识别为具有社会性意图(Yin et al., 2013)。这类研究提示, 除了行为的耦合性, 人们也可能基于其它线索或信息对社会性意图进行识别(Auvray et al., 2009)。因此, 本研究将进一步考察这一识别过程中可被利用的其他线索或信息。
社会性意图虽与客体导向性意图所指向的对象各异, 但其执行行为的主体均是存在理性的智能个体, 故也可基于动作效用最大化这一理性原则来分析交互意图的识别问题(Gergely & Jacob, 2012)。社会交互行为指向的对象是另一理性的社会个体, 只分析某个个体的动作效用也许不适用于这类存在多个行为主体的情境。研究表明人们以整体的方式对交互行为进行表征, 该表征方式暗示视觉系统可能在整体水平上对交互行为的效用最大化进行计算, 也即对交互多方行为的收益和成本进行综合考虑(Török et al., 2019)。据此, 当满足如下条件时:智能体A协助智能体B达成目标状态所需要的成本小于智能体B单独达成该目标状态所需要的成本时(简称为成本最小化, 如递送他人较难拿到的物品), 其达到整体层面的效用最大化, 可推测该动作模式被识别为具有社会性意图。也即, 视觉系统在识别社会性意图时, 可利用的线索包括成本最小化信息。但该假设并不排除社会性意图识别中存在其他线索的可能性, 如上文提到的耦合性, 仅假定成本最小化信息是社会性意图识别的充分而非必要条件。
本文中将采用脑电指标μ节律(8~13 Hz)对是否识别到社会性意图进行间接测量(如图1所示)。μ节律位于感觉运动皮层(C3和C4电极), 被认为产生于负责动作意图加工的重要功能区——镜像神经系统(Mirror Neuron System), 其中镜像神经系统活动越强, μ抑制越大(Muthukumaraswamy et al., 2004; Oberman et al., 2005)。研究发现, 由于社会性意图加工需同时考虑两个主体的行为, 该加工相比只包含单个个体的非社会性意图的加工, 对镜像神经系统的需求更高, 因此激活也更强(Centelles et al., 2011), 而镜像神经系统的活动可体现在μ节律的抑制上, 因此μ抑制的强度可用来指示被试是否将观察到的行为识别为社会性意图(Oberman et al., 2007; Yin et al., 2020)。其中, 相比非社会性的客体导向意图行为, 若某行为产生更强的μ抑制, 则可说明其被识别为具有更强的社会性意图, 且该指标已成功应用于社会性意图识别的相关研究(Oberman et al., 2007; Yin et al., 2020)。
图1
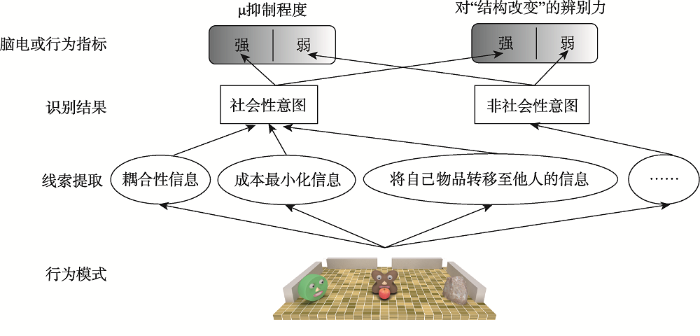
为检验上述假设, 本文借鉴Heider和Simmel (1944)的方法以动画来描绘不同的行为模式(如图2所示的刺激场景), 其行为主体均简化为几何形状式的智能体, 可用来模拟真实的交互场景且方便控制行为所引入的动作成本(Jara-Ettinger et al., 2015; Ullman et al., 2009)。本研究在动画中始终将智能体B保持静止, 而智能体A转移目标物苹果, 通过在B面前设置障碍物(即B是否被栅栏围住)的方法操纵B获得目标物苹果的成本。如此, 若A将目标物苹果放置在B前面, 由于智能体面前的物体被理解为归其所有(Tatone et al., 2015), 即B获得目标物苹果。当B前面存在栅栏时, 由于其难以逾越该障碍物, 此时A将该目标物放置在B面前的动作成本, 小于B通过自身的动作获得该目标物的成本; 但当B前面不存在栅栏时, 则B通过自身的动作获得该目标物的成本(路径长度)小于A将该目标物放置在B面前的动作成本。由于存在栅栏时满足成本最小化条件, 则该情境中A应被识别为具有社会性意图; 而对不存在栅栏的条件, 则被识别为非社会性的客体导向性意图或比存在栅栏时更弱的社会性意图。由于上述两种情境在物理层面存在差异(有无栅栏), 直接对比所发现的μ抑制差异可被物理层面的影响所解释。因此, 将上述两种实验分别与控制条件进行比较, 其被设置为在相同的情境中A采用同样的运动方式但将苹果放置在被栅栏围住的石头面前, 该行为与B无关, 属于典型的非社会性意图的行为模式(Tatone et al., 2015; Yin et al., 2020)。若人们确实可利用成本最小化信息这一线索识别社会性意图, 则当B被栅栏围住时, A将苹果放置在B面前(符合成本最小化)将被识别为具有社会性意图, 其μ抑制强于控制条件——A将苹果放置在石头前(实验1)。但当B没有被栅栏围住时, 无论A将苹果放置在B面前与否, 均不符合成本最小化条件, 因此A将苹果放置在B面前被识别为非社会性的客体导向意图或更弱的社会性意图, 可预测μ抑制程度与控制条件下无差异或差异小于有栅栏的情境(实验2)。
图2

图2
存在栅栏(a)和不存在栅栏(b)时, 智能体A将苹果放置在智能体B前或智能体B通过自身动作获得苹果的路径。
注:紫色实线为A的运动路径, 蓝色虚线为B的运动路径, 其趋近无穷大; 橙色实线为A的运动路径, 绿色虚线为B的运动路径。
为进一步检验本研究提出的假设, 本研究还采用了行为指标(如图1所示, 检测被试对不同变化的敏感性, 即辨别力)考察被试所识别的意图是否为社会性意图。实验采用变化觉察范式, 要求被试记忆一系列相同动作模式的动画(记忆项), 之后要求被试判断检测阶段的动画(检测项)是否在记忆项中出现过(如图7所示)。其中, 检测项的变化包括角色交换(即A将苹果放置在智能体B前变成B将苹果放置在智能体A前)、结构改变(即A将苹果放置在智能体B前和C将苹果放置在智能体D前, 变为C将苹果放置在智能体B前)。已有研究证实, 与格式塔组织相似, 两个存在社会交互意图的个体在工作记忆中会被组织在一起, 并作为整体单元分组进行存储(Stahl & Feigenson, 2014; Vestner et al., 2019; Ding et al., 2017)。鉴于记忆分组可提升被试对跨整体结构间的区分度, 但使得结构内的元素容易混淆(Papeo, 2020; Sedikides et al., 1993; Sherman et al., 2002; Suzuki & Cavanagh, 1995), 可推测当智能体A与B有交互意图时, 相比无交互意图时, 由于A与B被整合成一个具有交互关系的记忆分组, 人们更容易辨别其结构改变, 而对角色交换不敏感, 呈现记忆混淆效应(实验3a); 反之, 则辨别力相当(实验3b)。
2 实验1:存在成本最小化信息时的脑电频谱
本实验以顶叶的μ节律作为指标, 呈现智能体A将苹果放置在被栅栏围住的智能体B前面, 或是被栅栏围住的石头前面, 考察前者是否被识别为具有社会性意图。
2.1 被试
21名在校大学生自愿参与本实验, 其中1名被试因数据收集过程中电极脱落或运动幅度过大, 导致脑电数据质量较差, 故予以剔除。最终获得20名(其中男性8名, 女性12名)有效被试的实验数据, 被试的年龄在17~24岁之间(M = 19.5, SE = 0.4), 视力或矫正视力正常, 无色盲, 均无身心疾病。实验结束后, 被试获得50元人民币作为报酬。该样本量参考了以往用μ指标来探讨动作加工, 尤其是对交互行为加工的相关研究, 其样本量在20名被试左右, 结果的效应量处于中等水平(f介于0.25~0.46之间; Kourtis et al., 2019; Yin et al., 2017)。为保持实验样本量的一致性, 后续实验中也招募了20名有效被试。该实验获得了宁波大学心理学研究伦理委员会同意, 且在实验前保证每个被试充分知晓实验要求, 并签署了知情同意书。
2.2 实验仪器与刺激
实验采用19英寸CRT示器, 分辨率为800× 600, 刷新率为100 Hz, 被试距屏幕约60 cm。所有刺激材料的呈现和实验流程通过MATLAB的工具箱Psychophysics Toolbox (Brainard, 1997)来控制。
实验刺激材料为电脑动画, 其通过3D动画建模软件blender 2.78a制作而成, 显示大小为11.1°×8.3°。所有动画均设置为从12.2°的观看距离和73°的3D透视图的视角呈现。动画时长2 s, 其场景中包括两个不同的动画角色(一个运动的智能体, 以下称为“智能体A”, 与一个静止的智能体, 以下称为“智能体B”), 一个石头以及一个苹果。其中, 每个动画中智能体的外形相异, 且每个试次从36对不同的组合中随机选择(9种不同外形的智能体, 两两组合而得)。动画中的石头和智能体B分布在场景的两侧, 其距离中心2.8°, 且均被难以逾越的栅栏围着, 但在栅栏中间有个小洞口(智能体无法穿过), 以用于智能体A放置苹果。动画开始前苹果位于石头和智能体B的正中间, 智能体A位于屏幕中心上方1.6°处。整个动画中只有智能体A运动, A首先垂直向下运动至苹果所在位置(0.5 s), 之后向左或向右转向智能体B或石头(0.1 s), 接着向智能体B或石头运动, 并将苹果放置在B前方(0.5 s), 最后返回其初始位置(0.9 s)。根据苹果被放置目的地是智能体(图3a)还是石头(图3b), 分为两种实验条件。这两种条件除了智能体A放置苹果的目的地不同外, 具有相同的运动学特性。但当智能体A将苹果放置在智能体B前面时, 由于其动作成本小于B通过自身的动作获得该目标物的成本(需绕过几乎无法逾越的栅栏), 故符合成本最小化条件, 应被识别为具有社会性意图。实验动画示例可见
图3
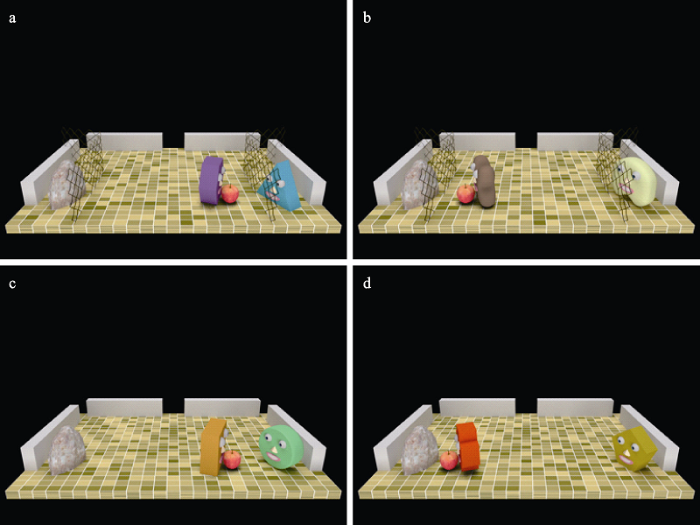
除了上述两种动画, 实验中还构建了其它动画, 作为填充动画, 要求被试计数其数量, 以使得被试的注意集中在屏幕的动画上。该类动画与上述动画基本相同, 除了在0.3~1.7 s之间智能体A会随机且连续消失0.8 s。
2.3 实验流程
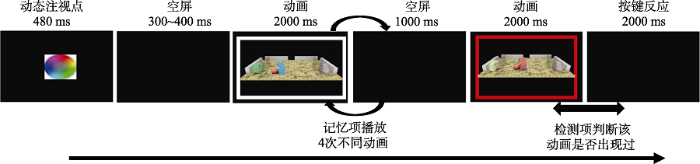
每个试次的流程如图4所示。首先, 在屏幕中央呈现一个动态的注视点, 持续480 ms。之后, 空屏300~400 ms。接着, 呈现动画的第一帧。300 ms后, 动画开始播放, 持续2 s。试次间隔为900~ 1100 ms。被试被要求认真观看屏幕中的动画, 并计数填充动画的次数, 在每个组块结束后进行报告。
图4

实验共有两个组块, 每个组块包含72个正式试次, 其中每个条件包含36个试次。每个条件36个试次动画中的智能体组合均不相同, 但条件间的智能体组合均相同。如此, 不同条件间除了情境, 其它的物理信息均一致。在前述设置的基础上, 每个组块还包括8~12个填充动画, 其用于注意检测。如果被试所报告的填充动画个数与真实的不一致, 则说明该被试可能未认真观看所有的视频。根据该注意检测标准, 被试均能准确报告填充动画个数。
2.4 数据记录
基于NeuroScan Synamps 2系统(Compumedics NeuroScan Inc.), 利用国际通用的10~20的64导Ag/AgCl电极帽记录EEG。以左侧乳突电极为参考。接地电极位于FPZ与FZ中央。以左眼眶额上、下部的2个电极记录垂直眼电(VEOG), 以两眼外侧1.5 cm处的2个电极记录水平眼电(HEOG)。脑电信号采集增益为500, 采样频率为500 Hz, 滤波带通(band-pass)为0.05~100 Hz。电极和头皮间的电阻均小于5 kΩ。
2.5 数据分析
采用MATLAB的EEGLAB和Fieldtrip工具包对数据进行分析。首先用EEGLAB对数据进行预处理。预处理中, 先将左侧乳突参考转换成左右乳突平均做参考, 之后对数据进行0.1~100 Hz的滤波(fir1)。接下来, 删除记录眼电的电极数据, 并进行独立成分分析(ICA)。对获得的独立成分, 采用EEGLAB的ADJUSR插件识别眼电成分与噪音成分, 并将其剔除1(1笔者也采用传统的分析方法剔除噪音试次, 即波幅超过±100 μV的试次被剔除, 在此基础上对μ频段能量进行分析, 其结果与采用ADJUSR插件删除噪音法所得结果相同)。。最后, 对脑电数据进行分段, 其长度为开始运动前500 ms到运动后2000 ms (动画结束点)。完成前述处理后, 采用Fieldtrip进行频谱分析。具体而言, 对每个分段数据, 首先就4~30 Hz的节律, 以0.5 Hz为步长进行Morlet小波分析(wavelet), 其带宽为3 Hz; 之后, 以运动开始前500 ms的能量值作为基线(此时间长度可保证包含目标分析频段至少3个完整的波长), 对每个分段数据(即开始运动前500 ms至开始运动后2000 ms)的能量值进行基线矫正; 最后, 对每个被试按条件进行平均。
与已有研究一致, 本实验选取的μ频段为8~ 13 Hz (Fox et al., 2016; Muthukumaraswamy et al., 2004; Ulloa & Pineda, 2007)。大量研究表明, 与动作加工相关的μ波段主要出现在顶叶运动区, 在脑电研究中常常选取顶叶的C3和C4电极(Duan et al., 2018; Pomiechowska & Csibra, 2017), 因此本实验也选取C3和C4电极对μ频段的激活模式进行分析。鉴于每个被试由于头皮厚度不同以及电阻不同可能导致μ频段激活的差异(Pineda & Oberman, 2006), 借鉴已有研究(Cuevas et al., 2014; Pfurtscheller & Lopes da Silva, 1999), 将动画呈现阶段的μ频段能量值减去基线的能量值并除以基线的能量值再乘100%作为μ的能量值。即对于每个被试, 因变量= (动画呈现阶段的μ频段能量-基线阶段的μ频段能量)/基线阶段的μ频段能量×100%, 其值小于零表示抑制。为进一步检验μ频段是否属于枕叶α活动的泛化, 参考以往研究(Klimesch et al., 2007; Perry et al., 2011), 本实验对枕叶O1和O2电极上的α也予以类似分析, 结果发现两个实验中不同类型动画所诱发的枕叶α活动并无差异(详见网络版附录, 其可在
在开展本文所报告的被试间设计实验前, 笔者也采用被试内设计收集了10个被试(结果见附录)。但每个被试均报告有无栅栏情境对比明显, 实验者设置栅栏是有目的的, 因此调整成了被试间设计。更重要的是, 对于智能体A将苹果放置在智能体B前, 有无栅栏情境在物理层面存在差异(栅栏的遮挡不同), 直接对比所发现的μ抑制差异可被物理层面的影响所解释。因此, 将是否存在栅栏时智能体A将苹果放置在智能体B前分别与控制条件进行比较, 即目前的被试间设计。
2.6 结果与讨论
不同条件下在不同电极上诱发的8~13 Hz频段能量及其差异地形图如图5和6所示。对μ相对能量值进行2 (电极:C3与C4) × 2 (放置目的地:智能体与石头)的重复测量方差分析, 结果发现:仅有放置目的地的主效应显著, F(1, 19) = 8.71, p = 0.008, η2 p= 0.31, 差异的95% CI = [-15.1%, -2.6%], 其中当动态智能体将苹果放置在静态智能体(智能体B)前时, μ频段能量(M = -17.3%, SE = 3.6%)显著强于(抑制程度更高)将其放置在静态石头前的μ频段能量(M = -8.5%, SE = 4.6%); 电极的主效应不显著, F(1, 19) = 0.80, p = 0.382; 电极与放置目的地的交互作用不显著, F(1, 19) < 0.01, p = 0.975。
图5
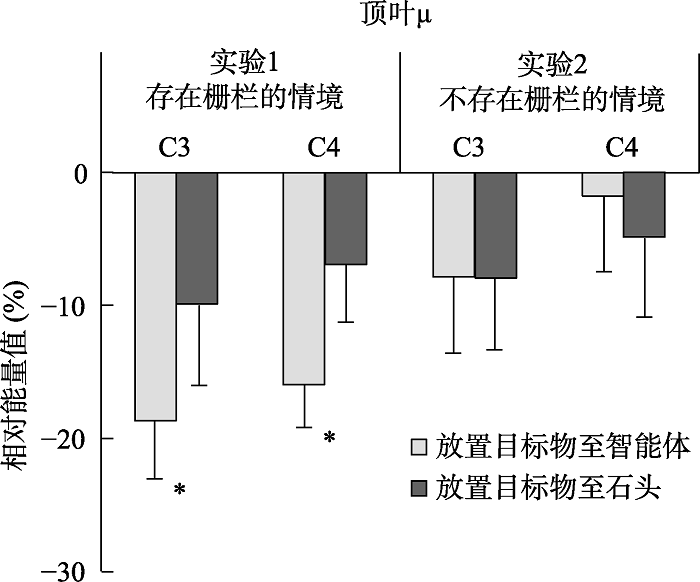
图6
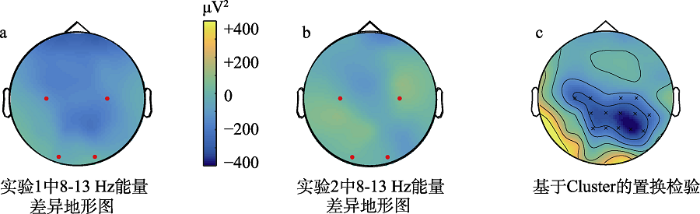
图6
有无栅栏情境下实验1(a)和实验2(b)中8~13 Hz频段的差异地形图以及(c)基于Cluster置换检验的地形图
注:c图中符号×表示存在显著差异的电极。
上述结果发现, 当智能体B被栅栏围住时, 智能体A将苹果放置在智能体B前时μ抑制强于控制条件——智能体A将苹果放置在石头前, 且并非α活动的泛化, 其与预测一致, 支持成本最小化信息是社会性意图识别重要线索的观点。
3 实验2:不存在成本最小化信息时的脑电频谱
实验1中当智能体A将苹果放置在智能体B前时, 智能体A的动作可减少智能体B完成目标的动作成本, 即符合成本最小化条件, μ的抑制程度显著高于被识别为客体导向性意图的控制条件。针对该结果, 可能也仅仅是一个智能体趋近另一个智能体, 而非成本最小化信息, 导致其被识别为社会性意图。本实验将通过去除成本最小化信息的方法, 对这一可能性进行检验。实验中, 智能体A与实验1保持相同的运动特性, 但将实验1中围住智能体B和石头的栅栏移除, 使得智能体B获得苹果的成本小于智能体A将苹果放置在其前面的成本(即不符合成本最小化)。若人们确实基于成本最小化信息这一线索识别社会性意图, 则当智能体A将苹果放置在未被栅栏围住的智能体B前时, 即不符合成本最小化, 实验1的效应消失; 反之, 若效应依然存在, 则说明智能体之间社会性意图识别取决于其它线索, 如:趋近智能体的行为。
3.1 方法
20名在校大学生自愿参与本实验(其中男性8名, 女性12名)。被试的年龄在17~24岁之间(M = 19.9, SE = 0.4), 视力或矫正视力正常, 无色盲, 均无身心疾病。
3.2 结果
为进一步检验实验1的结果确实由于动作识别的差异所致, 合并C3和C4电极上的数据, 形成2 (实验:实验1与实验2; 被试间变量) × 2(放置目的地:智能体与石头; 被试内变量)的实验设计, 对顶叶的μ频段能量进行方差分析。结果发现, 实验的主效应不显著, F(1, 38) = 1.33, p = 0.257; 放置目的地的主效应也不显著, F(1, 38) = 1.90, p = 0.176; 但两因素的交互作用显著, F(1, 38) = 4.27, p = 0.046, η2 p= 0.10。且就智能体A将苹果放置在智能体B前, 存在栅栏时所诱发的μ抑制显著(M = -17.3%, SE = 3.6%)强于不存在栅栏的情境(M = -4.8%, SE = 5.6%; t(38) = 1.89, p = 0.034, Cohen’s d = 0.60, 其为单尾检验), 而在智能体A将苹果放置在石头前时该差异消失(t(38) = 0.28, p = 0.390, Cohen’s d = 0.09)。该结果提示实验操纵调节不同动作模式下顶叶的μ频段能量, 进一步支持实验1的结果确实由于动作识别结果的不同所致。
4 实验3a:存在成本最小化信息时的行为反应
本实验呈现与实验1相同的动画, 但采用变化觉察范式, 基于信号检测论测量被试对不同变化的辨别力, 考察是否存在记忆混淆效应, 以进一步检验研究假设。
4.1 被试
保持与前述研究相同的样本量, 20名在校大学生自愿参与本实验(其中男性7名, 女性13名)。被试的年龄在17~24岁之间(M = 21.2, SE = 0.3), 视力或矫正视力正常, 无色盲, 均无身心疾病。
4.2 实验材料与流程
实验的刺激材料与实验1相同, 但流程改为变化觉察范式(如图7所示)。具体而言, 首先, 在屏幕中央呈现一个动态的注视点, 持续480 ms之后, 空屏300~400 ms。接着, 在白框中(11.1°× 8.3°)依次播放4个需要记忆的相同模式的动画作为记忆项, 每个持续2000 ms, 两者之间有1000 ms的空屏间隔。随后, 再呈现一个动画, 并伴随着白色边框变为红框以提醒被试在呈现检测项, 其持续2000 ms。在检测项消失后, 要求被试尽可能准确地判断检测项是否在记忆项中出现过。如果检测项与某个记忆项相同, 则要求被试按下“J”键; 否则, 按下“F”键。如果在2000 ms内未按键, 程序会自动进入下一个试次。每个试次间隔为1500 ms。
图7
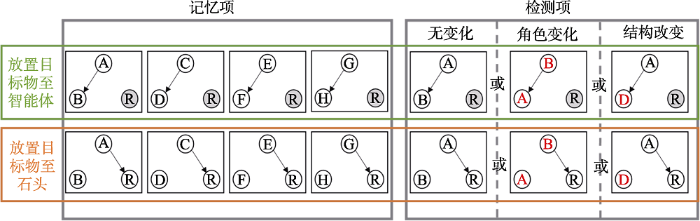
每个试次中, 所有记忆项和检测项的动作模式相同(即运动路径以及是将苹果放置在智能体前还是石头前), 但每个记忆项都是一对外观不同的智能体。在检测项中, 基于智能体外观的变化可区分为三种条件, 如图8所示:(1)无变化, 呈现与某个记忆项相同的一对智能体, 且智能体的角色保持一致(也即随机抽取一个记忆项作为检测项); (2)角色交换, 检测项中智能体来自某个记忆项, 但智能体的角色互换(即A将苹果放置在智能体B前变成B将苹果放置在智能体A前); (3)结构改变, 记忆项中某两个动画中占据相同角色的智能体互换(即A将苹果放置在智能体B前和C将苹果放置在智能体D前, 变为C将苹果放置在智能体B前)。据此, 由于记忆和检测项的动作模式相同, 被试只需根据动画中两个智能体的外观来判断动画的变化, 从而可以检测被试对动画中的两个智能体是否进行了记忆分组。
图8
为平衡每种变化类型下的试次数量, 无变化条件下的试次等于角色交换和结构改变条件的试次之和。对于无变化条件下的试次, 一半的试次被视为没有角色交换, 另一半被视为没有结构改变。事实上, 在无角色交换和无结构改变条件之间没有区别, 只是作为后续数据分析的区分。上述变化条件均出现在目标物放置在智能体前或石头前这两个条件下。每个条件有20个试次, 结果总共160个试次, 共分为4组, 每组结束后休息5分钟。所有试次以伪随机顺序呈现, 在连续三次试次中不重复相同的条件。
4.3 数据分析
为检验记忆混淆效应, 即放置目的地(智能体与石头)是否会调节被试对不同变化的敏感性, 根据信号检测论, 以“检测项与所有记忆项不同” (即变化)作为信号和“检测项与某个记忆项相同” (即无变化)作为噪音, 计算不同变化类型下的敏感性指标d’, 即辨别力(Yin et al., 2018)。为了避免在计算d’时出现无限大的情况, 参考Snodgrass和Corwi (1988)的建议, 对每种变化类型下所统计击中、漏报、虚报和正确拒斥的频次均加0.5, 且除以N+1。其中, N为对应条件下频次。据此, 可获得2 (变化类型:角色交换与结构改变) × 2 (放置目的地:智能体与石头)的被试内设计的辨别力, 对其进行重复测量方差分析。此外, 对基于信号检测论计算所得的决策标准β也进行了报告, 详见网络版附录。
4.4 结果
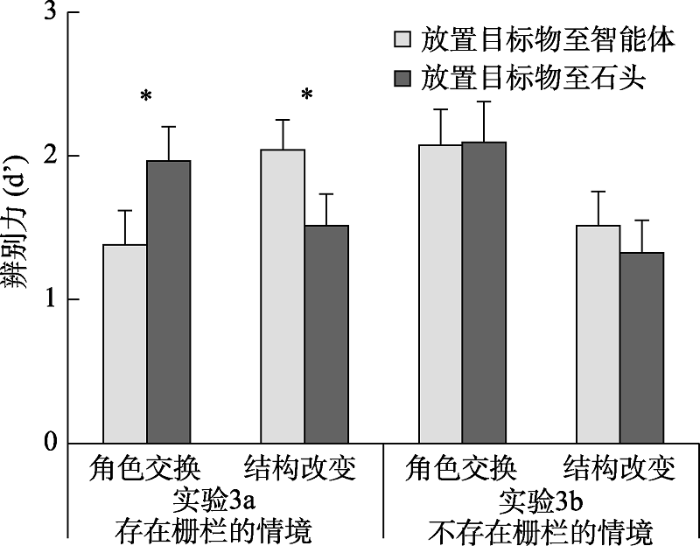
不同条件下的辨别力如图9所示。对辨别力进行2 (变化类型:角色交换与结构改变) × 2 (放置目的地:智能体与石头)的重复测量方差分析, 结果发现:变化类型的主效应不显著, F(1, 19) = 1.12, p =0.304; 放置目的地的主效应不显著, F(1, 19) = 0.11, p = 0.746; 但变化类型与放置目的地的交互作用显著, F(1, 19) = 8.03, p = 0.011, η2 p= 0.30。简单效应分析发现, 在放置目标物至智能体条件下, 角色交换(M = 1.38, SE = 0.24)的辨别力显著低于结构改变(M = 2.04, SE = 0.21; t(19) = 2.78, p = 0.012, Cohen’s d = 0.62), 而在放置目标物至石头条件时, 角色交换(M = 1.97, SE = 0.25)的辨别力显著高于结构改变(M = 1.51, SE = 0.23; t(19) = 2.26, p = 0.036, Cohen’s d = 0.50)。更重要的是, 在角色交换条件下, 被试对放置目的地为石头(M = 1.97, SE = 0.25)条件的辨别力显著高于放置目的地为智能体的条件(M = 1.38, SE = 0.24; t(19) = 2.33, p = 0.031, Cohen’s d = 0.52); 相反, 当变换类型为结构改变时, 放置目的地为智能体条件下(M = 2.04, SE = 0.21)辨别力显著高于放置目的地为石头的条件(M = 1.51, SE = 0.23; t(19) = 3.21, p = 0.005, Cohen’s d = 0.72), 呈现记忆混淆效应。
图9
5 实验3b:不存在成本最小化信息时的行为反应
为进一步检验成本最小化信息的作用, 与实验2相同, 实验3b将去除成本最小化信息, 观察实验3a的效应是否消失。
5.1 方法
20名在校大学生自愿参与本实验(其中男性7名, 女性13名)。被试的年龄在17~24岁之间(M = 20.1, SE = 0.4), 视力或矫正视力正常, 无色盲, 均无身心疾病。
实验设计和流程与实验3a基本相同, 除了趋近的目标物(智能体或石头)前面的栅栏不存在。数据分析与实验3a相同。
5.2 结果
不同条件下的辨别力如图9所示。对辨别力进行2 (变化类型:角色交换与结构改变) × 2 (放置目的地:智能体与石头)的重复测量方差分析, 结果发现:变化类型的主效应显著, F(1, 19) = 8.14, p = 0.010, η2 p= 0.30, 角色交换条件(M = 2.09, SE = 0.25)的辨别力显著高于结构改变(M = 1.42, SE = 0.22); 放置目的地的主效应不显著, F(1, 19) = 0.48, p = 0.495; 变化类型与放置目的地的交互作用不显著, F(1, 19) = 0.64, p = 0.434。
6 讨论
本研究针对视觉可利用哪些信息对社会性(交互)意图进行识别的问题, 提出当某个体A协助另一个体B达成目标状态所需要的成本小于个体B单独达成目标状态所需要的成本(成本最小化信息)时, 视觉系统会将其识别为动作发出方对受益方存在以交互为目的的社会性意图。为检验该假设, 呈现智能体A将苹果放置在智能体B前面或石头前面的动画, 并通过在智能体B前面设置障碍物(即B是否被栅栏围住)的方法操纵成本最小化信息, 以四项实验探讨了成本最小化信息是否是社会性意图识别的线索。结果显示, 相比被识别为客体导向性意图的控制条件(即A将目标物苹果放置在石头前), 当A将目标物苹果放置在被栅栏挡住的B前, 其动作可减少B单独获取该苹果的动作成本, 即符合成本最小化条件时, μ的抑制程度更高(实验1), 且对结构改变(某两个动画中充当相同角色的智能体互换)的辨别力更强, 但对角色交换(某个动画中两个智能体的角色交换)的辨别力更弱(实验3a); 而当栅栏不存在时, 虽然A的运动路径与实验1相同, 但A将苹果放置在B前的成本大于B自身获取苹果的成本, 即不符合成本最小化条件, 条件间μ抑制的差异消失(实验2), 且对不同动作模式中变化的辨别相当(实验3b)。
针对导致μ抑制差异的可能原因, 本研究设置了严格的控制条件, 以确保其只能被意图识别结果的不同所解释。首先, 鉴于存在栅栏和不存在栅栏时智能体A将苹果放置在智能体B前的物理差异不同(栅栏的存在以及对智能体B的遮挡程度不同), μ的抑制程度差异可能反映了物理层面的差异, 因此实验中分别将其与各自的控制条件——智能体A将苹果放置在石头前进行对比。该设计一方面可更好的排除物理差异的解释:因被试所看到的差异仅是智能体A运动朝向的不同, 通过随机化设置可在条件间保持平衡, 故出现μ激活程度的差异只能由A将苹果放置在B前所蕴含的特殊信息所致; 另一方面, 控制条件智能体A将苹果放置在石头前属于典型的客体导向性意图, 可为实验条件——智能体A将苹果放置在智能体B前提供解释基线:若后者μ激活程度与控制条件相当, 则可说明其也被识别为客体导向性意图, 而后者μ激活程度高于控制条件, 则可说明其被识别为社会性意图。再者, 对于成本化最小化信息进行了直接操纵(实验1与实验2), 结果发现在不存在成本最小化信息(不存在栅栏)时, 实验条件——智能体A将苹果放置在智能体B前, 与控制条件——智能体A将苹果放置在石头前μ激活程度相当, 揭示了实验1条件间μ激活程度的差异确是由于意图识别结果的不同所致。这一结果不仅重复了Yin等(2020)发现的μ激活程度随着社会性意图的增强而提升的结果, 且进一步扩展了Yin等人的研究, 揭示了适用于社会性意图识别的一种新线索(Yin et al., 2020)。Yin等(2020)的研究中苹果是在智能体A的面前, 且无论是A转移苹果的成本还是B自己拿苹果的成本都相同, 而关键是智能体A“将自己的物品转移至他人”这一线索引发了社会性意图的识别, 而本研究并无“将自己的物品转移至他人”的动画设置, 而对成本最小化这一线索进行了操纵。
除采用脑电指标对感知到的意图类型进行考察, 本研究还基于变化觉察范式, 采用信号检测论测量被试对不同变化的敏感性。对于存在社会性意图的两个智能体的记忆, 已有研究证明其可被整合成一个记忆分组单元(Stahl & Feigenson, 2014; Vestner et al., 2019; Ding et al., 2017), 因此人们更容易辨别其结构改变, 而对角色交换不敏感, 呈现记忆混淆效应。这一记忆效应出现在了当智能体A将苹果放置在被栅栏围住的智能体B前, 提示其被识别为具有社会性意图, 进一步验证了基于脑电获得的结论。而当栅栏被去除后, 即去除成本最小化信息, 仅发现对角色交换的辨别力高于结构变化, 其与存在栅栏时对实验3a中控制条件不同变化的辨别相当。究其原因, 动画中的施动者(智能体A)是运动的, 而受动者(智能体B)保持静止, 当智能体间不存在社会性意图或社会性意图较弱时, 二者并未被记忆分组, 而是以单个智能体进行存储。如此, 当运动的智能体变为静止的智能体时, 物理状态发生了较大改变, 从而使得被试更容易辨别。然而, 这一现象在存在成本最小化信息的条件中(智能体A将苹果放置在被栅栏围住的智能体B前)弱化, 甚至反转, 进一步说明了其被识别为具有社会性意图, 从而产生记忆分组。
本研究发现某智能体将苹果放置在另一智能体前(不存在成本最小化信息), 其被识别为不具有社会性意图, 其似乎与生活经验相悖。经验上, 只要两个人之间发生接触, 就极有可能被解释为存在社会性意图。但该经验化解释往往具备一定的前提:双方都有行为上的外显反应, 或即使另一方无行为上的反应, 也会赋予内部心理的变化, 如:他内心嫉恨对方。但在人们的日常经验中所涉及的多是有外显反应的交互双方(如握手), 其存在社会性意图识别中的另一重要线索——耦合性线索, 所以会被解释为存在社会性意图。就第一个可能前提, 本研究中只有一方有行为反应(即运动), 而另一方永远静止, 因此很难满足。针对第二个可能前提, 实验2中智能体A将苹果放置在智能体B前确实可认为A想与B交互, 但B不愿意, 但这一解释强烈依赖于对他人的心智进行经验推测, 且强烈依赖于个人的经验, 不具有推理结果的稳定性, 因为不同人有不同的解释。其实, 深入分析也可发现将目标物放置在某人面前并不代表有社会性意图, 如:考试开始前学生将书本放在老师所在的讲台前。为何会产生不同层面的解释?这与人类对行为理解的认知系统有关。人类对行为理解的认知系统被认为分为心智化表征(mentalistic representation)和目的论表征(teleological representation; Gergely & Csibra, 2003)。前者基于对他人可能心理状态的推测, 以解释他人的动作, 其有赖于经验对心理状态的有意思考, 如:即使看到某人表面上对他人行为友善, 观察者可认为这个人内心想加害他人, 因此难以成为社会性意图推理中的通用线索, 其因人而异。而后者基于可被直接观察的要素, 如:情境的约束、动作的成本和收益等这些可容易观察到的要素, 基于效用最大化原则对他人的动作意图进行推测, 其具有自动化加工特性, 不需要任务引导, 其观察到的信息可成为人们识别动作的重要线索之一(Buon et al., 2013)。本研究中, 被试不需要有意对所观察到的动作进行推理, 而是被动地观察并计数可能的填充试次, 更多依赖于对动作的目的论表征。基于该表征, 检测到可依赖的线索——社会性意图识别的充分条件, 即成本最小化信息, 从而将其识别为具有社会性意图。但针对这一识别过程, 若成本最小化信息缺失, 人们可能还会搜寻其他可依赖的线索, 甚至受到个人对他人心智推测经验的影响, 后续研究将对这些问题进一步探讨。
本研究以动画的形式模拟动作, 可对动作发生的情境进行有效操纵, 且可对额外变量进行严格控制, 此方法已被广泛应用于动作理解领域。但该方法所设置的动作对人类的动作进行了一定程度抽象, 其发现的结论还有待在真正的人类动作上检验生态效度。同时, 为有效控制交互双方的动作成本, 本研究对成本最小化信息的操纵主要以智能体B实现目标的路径长短实现, 后续研究可进一步操作智能体B获取目标的路径长度, 并采用心理物理法获取成本和收益相同的主观等值点, 以此区分成本最小化信息的分割点, 从而检验本研究结论的普遍性。
7 结论
本研究通过设置障碍物的方法操纵当个体A协助另一个体B达成目标状态所需要的成本是否小于个体B单独实现该目标状态所需要的成本, 即是否存在成本最小化信息, 以指示不同意图类型的脑电μ以及对不同变化的辨别力为指标, 探讨了人们是否利用成本最小化信息对社会性意图进行识别, 获得以下结论:两个个体的行为是否满足成本最小化影响人们对动作意图的识别, 揭示成本最小化信息是识别社会性意图的重要线索。
附录:
1 枕叶α频段能量
1.1 实验1
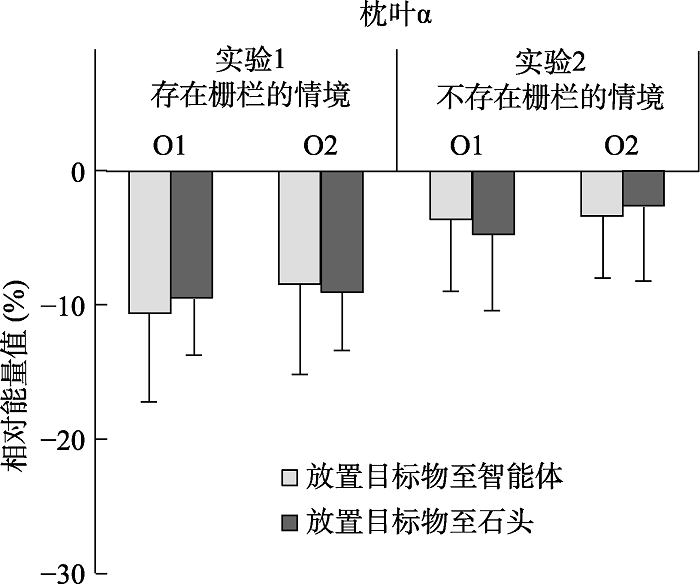
不同条件下枕叶α频段能量如附图1所示。以枕叶α频段能量作为因变量, 2(电极:O1与O2) × 2(放置目的地:智能体与石头)的重复测量方差分析结果发现:电极的主效应(F(1, 19) = 0.47, p = 0.500)、放置目的地的主效应(F(1, 19) = 0.01, p = 0.920)以及二者的交互作用(F(1, 19) = 0.67, p = 0.423)均不显著。该结果提示, 不同类型动画所诱发的μ能量差异并非枕叶α活动的泛化。
附图1
1.2 实验2
不同条件下枕叶α频段能量如附图1所示。同时, 就枕叶α频段能量而言, 2(电极:O1与O2) × 2(放置目的地:智能体与石头)的重复测量方差分析未发现任何效应显著。其中, 电极的主效应(F(1, 19) = 0.24, p = 0.628)、放置目的地的主效应(F(1, 19) < 0.01, p = 0.957)以及二者的交互作用(F(1, 19) = 0.97, p = 0.337)均不显著。
2 决策标准
2.1 实验3a
就决策标准(附图2)而言, 2(变化类型:角色交换与结构改变) × 2(放置目的地:智能体与石头)的重复测量方差分析并未发现任何效应显著。其中, 变化类型的主效应(F(1, 19) = 3.60, p = 0.073)、放置目的地的主效应(F(1, 19) < 0.01, p = 0.953)以及二者的交互作用(F(1, 19) = 0.13, p = 0.718)均不显著。
附图2
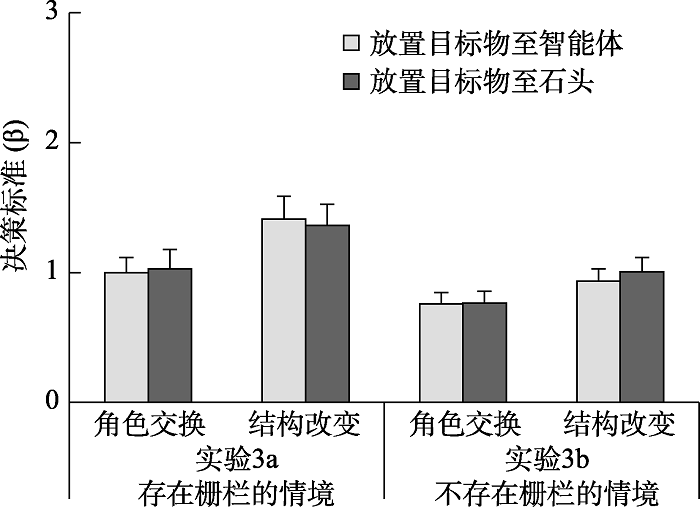
2.2 实验3b
就决策标准(附图2)而言, 2(变化类型:角色交换与结构改变) × 2(放置目的地:智能体与石头)的重复测量方差分析未发现任何效应显著。其中, 变化类型的主效应(F(1, 19) = 3.24, p = 0.088)、放置目的地的主效应(F(1, 19) = 0.26, p = 0.618)以及二者的交互作用(F(1, 19) = 0.16, p = 0.693)均不显著。
3 被试内设计初步结果
关于被试内设计, 初步收集了10名被试。结果发现, 存在栅栏时, 放置至智能体的激活显著强于放置至石头(Wilcoxon W = 5.00, p = 0.020, Cohen’s d = 0.82); 而不存在栅栏时, 激活不存在显著差异(Wilcoxon W = 34.00, p = 0.557, Cohen’s d = 0.24), 且当智能体A将苹果放置至智能体B时, 存在栅栏时的激活存在显著强于不存在栅栏时激活的趋势(Wilcoxon W = 9.00, p = 0.064, Cohen’s d = 0.67)。
参考文献
Perceptual interactions in a minimalist virtual environment
DOI:10.1016/j.newideapsych.2007.12.002 URL [本文引用: 1]
From the perception of action to the understanding of intention
The psychophysics toolbox
The Psychophysics Toolbox is a software package that supports visual psychophysics. Its routines provide an interface between a high-level interpreted language (MATLAB on the Macintosh) and the video display hardware. A set of example programs is included with the Toolbox distribution.
A non- mentalistic cause-based heuristic in human social evaluations
DOI:10.1016/j.cognition.2012.09.006 URL [本文引用: 1]
The neural bases of social intention understanding: The role of interaction goals
DOI:10.1371/journal.pone.0042347 URL [本文引用: 1]
Recruitment of both the mirror and the mentalizing networks when observing social interactions depicted by point-lights: A neuroimaging study
DOI:10.1371/journal.pone.0015749 URL [本文引用: 1]
One- year-old infants use teleological representations of actions productively
DOI:10.1207/s15516709cog2701_4 URL [本文引用: 1]
The infant EEG mu rhythm: Methodological considerations and best practices
DOI:10.1016/j.dr.2013.12.001 URL [本文引用: 1]
Can social interaction constitute social cognition?
DOI:10.1016/j.tics.2010.06.009
PMID:20674467
[本文引用: 1]

An important shift is taking place in social cognition research, away from a focus on the individual mind and toward embodied and participatory aspects of social understanding. Empirical results already imply that social cognition is not reducible to the workings of individual cognitive mechanisms. To galvanize this interactive turn, we provide an operational definition of social interaction and distinguish the different explanatory roles - contextual, enabling and constitutive - it can play in social cognition. We show that interactive processes are more than a context for social cognition: they can complement and even replace individual mechanisms. This new explanatory power of social interaction can push the field forward by expanding the possibilities of scientific explanation beyond the individual.Copyright © 2010 Elsevier Ltd. All rights reserved.
Two equals one: Two human actions during social interaction are grouped as one unit in working memory
DOI:10.1177/0956797617707318 URL [本文引用: 2]
Automatic attribution of social coordination information to chasing scenes: Evidence from mu suppression
DOI:10.1007/s00221-017-5111-4 URL [本文引用: 1]
Assessing human mirror activity with EEG mu rhythm: A meta-analysis
DOI:10.1037/bul0000031 URL [本文引用: 1]
Teleological reasoning in infancy: The naïve theory of rational action
Converging evidence demonstrates that one-year-olds interpret and draw inferences about other's goal-directed actions. We contrast alternative theories about how this early competence relates to our ability to attribute mental states to others. We propose that one-year-olds apply a non-mentalistic interpretational system, the 'teleological stance' to represent actions by relating relevant aspects of reality (action, goal-state and situational constraints) through the principle of rational action, which assumes that actions function to realize goal-states by the most efficient means available. We argue that this early inferential principle is identical to the rationality principle of the mentalistic stance - a representational system that develops later to guide inferences about mental states.
Reasoning about instrumental and communicative agency in human infancy
An experimental study of apparent behavior
DOI:10.2307/1416950 URL [本文引用: 1]
Goal attributions and instrumental helping at 14 and 24 months of age
DOI:10.1016/j.cognition.2015.03.014 URL [本文引用: 1]
Children’s understanding of the costs and rewards underlying rational action
DOI:10.1016/j.cognition.2015.03.006
PMID:25867996
[本文引用: 2]
Humans explain and predict other agents' behavior using mental state concepts, such as beliefs and desires. Computational and developmental evidence suggest that such inferences are enabled by a principle of rational action: the expectation that agents act efficiently, within situational constraints, to achieve their goals. Here we propose that the expectation of rational action is instantiated by a naïve utility calculus sensitive to both agent-constant and agent-specific aspects of costs and rewards associated with actions. In four experiments, we show that, given an agent's choices, children (range: 5-6 year olds; N=96) can infer unobservable aspects of costs (differences in agents' competence) from information about subjective differences in rewards (differences in agents' preferences) and vice versa. Moreover, children can design informative experiments on both objects and agents to infer unobservable constraints on agents' actions.Copyright © 2015 Elsevier B.V. All rights reserved.
Event-related phase reorganization may explain evoked neural dynamics
DOI:10.1016/j.neubiorev.2007.03.005 URL [本文引用: 1]
Evidence for we-representations during joint action planning
DOI:10.1016/j.neuropsychologia.2019.05.029 URL [本文引用: 1]
Nonparametric statistical testing of EEG- and MEG-data
In this paper, we show how ElectroEncephaloGraphic (EEG) and MagnetoEncephaloGraphic (MEG) data can be analyzed statistically using nonparametric techniques. Nonparametric statistical tests offer complete freedom to the user with respect to the test statistic by means of which the experimental conditions are compared. This freedom provides a straightforward way to solve the multiple comparisons problem (MCP) and it allows to incorporate biophysically motivated constraints in the test statistic, which may drastically increase the sensitivity of the statistical test. The paper is written for two audiences: (1) empirical neuroscientists looking for the most appropriate data analysis method, and (2) methodologists interested in the theoretical concepts behind nonparametric statistical tests. For the empirical neuroscientist, a large part of the paper is written in a tutorial-like fashion, enabling neuroscientists to construct their own statistical test, maximizing the sensitivity to the expected effect. And for the methodologist, it is explained why the nonparametric test is formally correct. This means that we formulate a null hypothesis (identical probability distribution in the different experimental conditions) and show that the nonparametric test controls the false alarm rate under this null hypothesis.
Mu rhythm modulation during observation of an object-directed grasp
Recent electrophysiological studies have shown that the human electroencephalographic mu rhythm is suppressed during the observation of actions performed by other persons, an effect that may be functionally related to the behaviour of so-called "mirror neurons" observed in area F5 of nonhuman primates. Because mirror neuron activity has been reported to be functionally specific to object-oriented actions, the present study was designed to determine if the human mu rhythm also exhibits this property. EEG measurements were obtained from 12 normal subjects while they observed either a precision grip of a manipulandum or an empty grip using the same hand position. Our results showed that the magnitude of the mu rhythm was significantly lower for the object grip condition than for the empty grip condition. These data support the notion that the human mu rhythm indexes a brain system that is functionally comparable to the monkey mirror neuron system. We propose that nonobject-directed actions may result in representational schemas that are either different or less salient than motorically equivalent actions that are directed toward objects.
EEG evidence for mirror neuron dysfunction in autism spectrum disorders
DOI:10.1016/j.cogbrainres.2005.01.014 URL [本文引用: 1]
The human mirror neuron system: A link between action observation and social skills
Twos in human visual perception
DOI:10.1016/j.cortex.2020.06.005 URL [本文引用: 1]
Motor and attentional mechanisms involved in social interaction-evidence from mu and alpha EEG suppression
DOI:10.1016/j.neuroimage.2011.06.060
PMID:21742042
[本文引用: 1]
Mu rhythms are EEG oscillations in the 8-13 Hz recorded at sites located roughly over the sensory-motor cortex. There is reliable evidence that the amplitude of mu rhythms is reduced when the participant performs a motor act (mu suppression). Recent studies found mu suppression not only in response to actual movements but also while the participant observes actions executed by someone else. This finding putatively associates the mu suppression to the activity of a mirror neurons system which, in humans, has been suggested to contribute to social skills. In the present study we explored the effects of different levels of social interaction on mu suppression. Participants observed dynamic displays of hand gestures performing actions used in the Rock-Scissors-Paper game. In different blocks, participants passively viewed identical video clips with no game context and in the context of a game, or while being actually engaged in the game either by imagining actions or by actual playing. As a baseline for calculating mu suppression we used a dynamic display of a rolling ball. In addition, to isolate the social aspect of the actual movements, participants performed the same acts outside the game context. Mu suppression was larger while participants were engaged in the social game than when they passively looked at the "opponent" actions or when they performed movements without the game context. This effect was found while viewing the opponent play as well as while actually playing, which supports the view that mu suppression is affected not only by motion, but also by the social context of the motion. However, we did not find differences in mu suppression between perception segments in which the participant did not actually play. Furthermore, in all perception segments occipital alpha suppression was more robust than mu suppression suggesting the involvement of a strong attentional component. While actually playing, however, mu suppression was stronger than alpha suppression.Copyright © 2011 Elsevier Inc. All rights reserved.
Event- related EEG/MEG synchronization and desynchronization: Basic principles
An internally or externally paced event results not only in the generation of an event-related potential (ERP) but also in a change in the ongoing EEG/MEG in form of an event-related desynchronization (ERD) or event-related synchronization (ERS). The ERP on the one side and the ERD/ERS on the other side are different responses of neuronal structures in the brain. While the former is phase-locked, the latter is not phase-locked to the event. The most important difference between both phenomena is that the ERD/ERS is highly frequency band-specific, whereby either the same or different locations on the scalp can display ERD and ERS simultaneously. Quantification of ERD/ERS in time and space is demonstrated on data from a number of movement experiments.
What goads cigarette smokers to smoke? Neural adaptation and the mirror neuron system
One model of addiction suggests that neural circuits in the frontal cortex adapt to drug use and become sensitized leading to excessive attribution of incentive salience to drug-associated cues. The present study examined changes associated with cigarette use in the frontal mirror neuron system (MNS) of the human brain, as reflected in mu rhythm responsiveness. Mirror neurons in premotor cortex exhibit visuomotor properties that allow them to respond to self-movement as well as the observation of movement. This is a potential neural substrate for imitation learning and social cognition, factors that may be important in determining who does and does not develop addictive behaviors. EEG mu rhythm suppression is hypothesized to reflect MNS activity and thus provide a non-invasive method for studying this relationship. Our results show that while nonsmokers exhibit normal mu suppression to observed and self-generated actions, smokers exhibit normal suppression only to self-movement but not to the observation of movement, particularly actions involving addiction-related cues. Non-abstinent and abstinent smokers (those abstaining for approximately 12 h) did not differ significantly in their responses to the observation of movement, i.e., both exhibited atypical patterns of mu rhythm reactivity compared to nonsmokers. These data support the hypothesis that cigarette use produces short- and longer term adaptations in the MNS. Such adaptations may inappropriately bias attention toward motivationally salient, addiction-related cues leading to more impulsive and addiction-related behaviors.
Motor activation during action perception depends on action interpretation
DOI:S0028-3932(17)30039-8
PMID:28189494
[本文引用: 1]
Since the discovery of motor mirroring, the involvement of the motor system in action interpretation has been widely discussed. While some theories proposed that motor mirroring underlies human action understanding, others suggested that it is a corollary of action interpretation. We put these two accounts to the test by employing superficially similar actions that invite radically different interpretations of the underlying intentions. Using an action-observation task, we assessed motor activation (as indexed by the suppression of the EEG mu rhythm) in response to actions typically interpreted as instrumental (e.g., grasping) or referential (e.g., pointing) towards an object. Only the observation of instrumental actions resulted in enhanced mu suppression. In addition, the exposure to grasping actions failed to elicit mu suppression when they were preceded by speech, suggesting that the presence of communicative signals modulated the interpretation of the observed actions. These results suggest that the involvement of sensorimotor cortices during action processing is conditional on a particular (instrumental) action interpretation, and that action interpretation relies on inferential processes and top-down mechanisms that are implemented outside of the motor system.Copyright © 2017 Elsevier Ltd. All rights reserved.
Relationships as natural categories
DOI:10.1037/0022-3514.64.1.71 URL [本文引用: 1]
The spontaneous use of a group typology as an organizing principle in memory
Five studies investigated the spontaneous use of group typology in encoding information about various social groups. Participants saw faces or behaviors along with a label indicating the group membership of the face or the behavior. Labels corresponded to 2 groups each of 3 group types (i.e., 2 intimacy groups, 2 task-oriented groups, and 2 social categories). Recognition results showed more within-group-type errors than between-group-types errors. A free-recall task replicated these results, as the sequence of remembering items showed that memory organization reflected the group typology. A final study investigated the effects of group typology on the speed and accuracy of category membership verification. Results demonstrate the spontaneous use of an implicit group typology and its influence on the cognitive organization of information about groups.
Pragmatics of measuring recognition memory: Applications to dementia and amnesia
DOI:10.1037/0096-3445.117.1.34 URL [本文引用: 1]
Social knowledge facilitates chunking in infancy
DOI:10.1111/cdev.2014.85.issue-4 URL [本文引用: 2]
Facial organization blocks access to low-level features: An object inferiority effect
DOI:10.1037/0096-1523.21.4.901 URL [本文引用: 1]
Giving and taking: Representational building blocks of active resource-transfer events in human infants
DOI:S0010-0277(14)00287-X
PMID:25614012
[本文引用: 2]
Active resource transfer is a pervasive and distinctive feature of human sociality. We hypothesized that humans possess an action schema of giving specific for representing social interactions based on material exchange, and specified the set of necessary assumptions about giving events that this action schema should be equipped with. We tested this proposal by investigating how 12-month-old infants interpret abstract resource-transfer events. Across eight looking-time studies using a violation-of-expectation paradigm we found that infants were able to distinguish between kinematically identical giving and taking actions. Despite the surface similarity between these two actions, only giving was represented as an object-mediated social interaction. While we found no evidence that infants expected the target of a giving or taking action to reciprocate, the present results suggest that infants interpret giving as an inherently social action, which they can possibly use to map social relations via observing resource-transfer episodes. Copyright © 2014 Elsevier B.V. All rights reserved.
Rationality in joint action: Maximizing coefficiency in coordination
DOI:10.1177/0956797619842550 URL [本文引用: 1]
Help or hinder: Bayesian models of social goal inference
Recognition of point-light biological motion: Mu rhythms and mirror neuron activity
DOI:10.1016/j.bbr.2007.06.007 URL [本文引用: 1]
Bound together: Social binding leads to faster processing, spatial distortion, and enhanced memory of interacting partners
DOI:10.1037/xge0000545 URL [本文引用: 2]
Social coordination information in dynamic chase modulates EEG Mu rhythm
DOI:10.1038/s41598-016-0028-x URL [本文引用: 1]
Social grouping: Perceptual grouping of objects by cooperative but not competitive relationships in dynamic chase
DOI:10.1016/j.cognition.2013.06.013 URL [本文引用: 1]
Giving, but not taking, actions are spontaneously represented as social interactions: Evidence from modulation of lower alpha oscillations
DOI:10.1016/j.neuropsychologia.2020.107363 URL [本文引用: 6]
Object- based attention on social units: Visual selection of hands performing a social interaction
DOI:10.1177/0956797617749636 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}