

1 问题提出
记忆是个体对其经验的识记、保持以及再认或回忆。过去经验对当前记忆的影响可以是有意识的, 也可以是无意识的。有意识的记忆称为外显记忆, 相对的, 无意识的记忆则称为内隐记忆。研究者对于这两种记忆之间的关系进行了大量的研究, 许多研究发现, 两种记忆在不同变量的影响下产生不一致的变化, 如加工水平(Alipour, Aerab-Sheybani, & Akhondy, 2012; 唐小庭, 2013)、干扰(孟迎芳, 于海莉, 2012; 林无忌, 孟迎芳, 林静远, 2017)、年龄 (Verneau, van der Kamp, Savelsbergh, & de Looze, 2014; Narme, Peretz, Strub, & Ergis, 2016)等。在认知神经科学的研究上, 也观察到两者的分离(Cabeza & Moscovitch, 2013; Korsnes & Magnussen, 2014)。虽然有大量研究证明内隐记忆与外显记忆在行为结果与神经机制上表现出分离, 但也有研究者认为, 这种分离现象是由于两者使用不同任务导致, 两种记忆实际从属于同一个记忆系统(Roediger & McDermott, 1993)。可见, 内隐记忆与外显记忆的分离现象是由于两种记忆任务不同, 还是两种记忆的生理机制不同仍然存在争议。
在使用不同变量研究各因素对记忆影响的实验中, 药物也是一个常见的变量, 其中胆碱相关药物又常在记忆研究中被使用。有研究发现, 胆碱类药物可以增强对记忆相关脑区的兴奋性输入, 并促进记忆编码(Buccafusco, Letchworth, Bencherif, & Lippiello, 2005; Levin, McClernon, & Rezvani, 2006; Kukolja, Thiel, & Fink, 2009)。关于胆碱如何对记忆产生影响, 主要有两种理论:特异性学说与非特异性学说。特异性学说认为, 记忆突触即胆碱突触, 胆碱神经通路本身参与记忆痕迹的形成。因此, 兴奋或抑制胆碱突触的药物可以直接作用于记忆痕迹形成过程, 从而易化或抑制信息的存储。而非特异性学说则认为, 胆碱系统主要通过影响与觉醒相关的神经, 而间接地影响记忆。引起人体内胆碱水平变化的药物分为拟胆碱药和抗胆碱药, 拟胆碱药能使人体内胆碱水平上升, 而抗胆碱药则具有与拟胆碱药相反的作用。研究发现, 胆碱相关药物的摄入能对记忆产生影响, 主要表现为摄入适量的拟胆碱药物后记忆成绩上升, 而摄入抗胆碱药物后记忆成绩下降(Bentley, Driver, & Dolan, 2009; Dumas et al., 2010; Rosier et al., 1999)。例如, 在Bentley等人(2009)的研究中, 被试在使用拟胆碱药物毒扁豆碱或安慰剂后进行图片再认任务。结果发现在使用拟胆碱药物后, 记忆成绩显著高于安慰剂组。而在Dumas等人(2010)的研究中, 被试在使用抗胆碱药物莨菪碱或安慰剂后进行词汇再认任务, 结果发现相对于安慰剂组, 被试在使用抗胆碱药物后降低了记忆成绩。
虽然有大量关于胆碱对记忆影响的研究, 但是这些研究主要探讨胆碱与外显记忆之间的联系, 对内隐记忆是否产生影响并不明确。有研究发现, 胆碱药物仅能对外显记忆产生影响, 而不影响内隐记忆(Danion et al., 1990; Knopman, 1991; Schifano & Curran, 1994)。Danion等人(1990)使用莨菪碱、三甲丙咪嗪和安定研究抗胆碱药物对内隐记忆与外显记忆的影响, 实验使用词干补笔与自由回忆任务测量内隐记忆与外显记忆。被试分别使用3种药物或安慰剂后进行记忆任务, 结果发现在自由回忆任务中, 使用3种药物的被试外显记忆成绩都低于安慰剂组。而在词干补笔任务中, 4组之间的内隐记忆成绩没有差异。Schifano和Curran (1994)使用氯羟去甲安定与莨菪碱对内隐与外显记忆进行研究, 结果同样发现药物损害了外显记忆, 但是对内隐记忆没有影响。但是, 也有研究认为胆碱药物不仅能对外显记忆产生作用, 同时也能对内隐记忆产生作用。在Bentley, Vuilleumier, Thiel, Driver和Dolan (2003)的研究中发现, 在面孔重复呈现时, 拟胆碱药物毒扁豆碱增强了次级枕叶皮层等区域的激活衰减现象, 而激活衰减现象是内隐记忆在脑成像结果上的表现。
胆碱药物可以影响外显记忆已经较为明确, 但是为何在对内隐记忆上的影响却有不同?我们推测不同结果的原因主要来源于几个方面。首先是实验任务, 在Danion等人(1990)与Schifano和Curran (1994)的研究中, 都使用词干补笔任务对内隐记忆进行测量。而在Bentley等人(2003)的研究中, 则通过比较重复刺激的激活衰减来代表内隐记忆。虽然两种任务都可以代表内隐记忆, 但同一变量对不同内隐记忆任务可能产生不同的影响。例如在提取干扰对内隐记忆的影响中, Lozito和Mulligan (2010)与Prull, Lawless, Marshall和Sherman (2016)使用词干补笔任务研究干扰对内隐记忆的影响时, 发现内隐记忆不受提取阶段干扰的影响, 而林无忌等人(2017)使用词汇判断任务则发现相反的结果。林无忌等人认为, 实验任务不同可能是造成不同结果的原因之一, 由于不同任务所要求的加工过程不同, 因此干扰影响了词汇判断的特定加工过程导致其成绩受到影响。其次, 虽然在Danion等人(1990)的研究中, 3组药物组的内隐记忆成绩与安慰剂组之间没有差异, 但是Danion在文中提到, 在莨菪碱组中发现内隐记忆的成绩与外显记忆成绩具有显著的相关, 而在安慰剂组中则没有这种相关。Danion认为两种记忆成绩在药物处理下呈现出显著相关, 可能说明两种记忆在胆碱药物的影响下有一定的关联, 因此并不能说胆碱对内隐记忆完全没有影响。最后, 实验中样本量较少, 上述实验中的被试量为10~15人, 较少的样本量可能造成统计结果不能反应真实情况。综上所述, 我们推测胆碱可能对内隐记忆产生一定影响。
在拟胆碱药物尼古丁对外显记忆影响的研究还发现, 尼古丁对记忆不同加工水平存在不同影响(Warburton, Skinner, & Martin, 2001; FitzGerald et al., 2008)。Warbuton认为尼古丁之所以对不同加工水平的影响不同, 是由于在对记忆项目进行语义加工时需要进行一定程度的联想。而尼古丁的摄入, 影响了语义加工的联想过程, 进而使记忆成绩发生变化。FitzGerald的研究进一步证实这种推论, 研究中发现由于语义和非语义加工所调用的脑区不同, 胆碱仅影响了语义加工所调用的脑区, 而没有影响非语义加工的脑区。这些研究都证明尼古丁对外显记忆不同加工水平的影响不同, 那么尼古丁对不同加工水平产生不同影响的现象是否会发生在内隐记忆中呢?
以往研究认为, 内隐记忆任务较不容易受到编码阶段加工水平的影响(Brooks, Gardiner, Kaminska, & Beavis, 2001; Rugg et al., 1998; Alipour et al., 2012)。例如, 在Brooks等人(2001)的研究中, 要求被试对名人姓名进行深浅加工, 然后进行内隐记忆与外显记忆测验。结果表明, 只有外显记忆测验结果表现出显著的加工水平效应, 而在内隐记忆中则没有发现。在认知神经方面的研究也发现相似的结果, 在Rugg等人(1998)的研究中发现, 加工水平对于外显记忆中的P600新旧效应影响较大, 但是对内隐记忆的N400新旧效应的影响则较小。内隐记忆不易受加工水平影响的现象与迁移适当加工理论较为符合, 根据该理论, 外显记忆依赖于概念加工, 而深加工是对记忆项目进行意义的判断, 更多地涉及概念层面的加工, 因此经过深加工后的项目能得到更好的外显记忆成绩。内隐记忆则依赖于知觉加工, 无论深浅加工, 记忆项目的知觉表征都被编码处理。但是内隐记忆与深加工水平中的概念加工无关, 经过概念加工的记忆项目并不能使内隐记忆成绩发生变化, 因此深浅加工之间的内隐记忆成绩没有差异。根据上述理论, 内隐记忆依赖于知觉加工, 不涉及概念加工, 而尼古丁对记忆产生的影响主要发生在语义加工所调用的脑区。如果该理论正确, 则可以推论尼古丁对内隐记忆的深浅加工的影响应该一致, 因此我们在实验中设置不同加工水平这一变量, 以进一步探索内隐记忆与外显记忆的关系。
此外, 胆碱对外显记忆不同阶段的影响并不相同(Rogers & Kesner, 2003; Gais & Born, 2004)。从信息加工的角度, 记忆可以分为编码、存储、提取三个阶段。在Rogers和Kesner (2003)与Gais和Born (2004)的研究中都发现, 拟胆碱药物能促进记忆的编码, 但是会损害提取。脑成像研究也发现, 拟胆碱药物使编码阶段的激活程度增强, 而使提取阶段的激活程度降低(Kukolja et al., 2009)。Kukolja认为这主要是由于编码与提取对胆碱水平的需要不同导致, 编码阶段较高的胆碱浓度能使编码过程更好地进行, 而提取则相反。胆碱对外显记忆的不同阶段能产生不同的影响, 那么胆碱对内隐记忆的不同阶段是否也能产生不同的影响呢?孟迎芳和郭春彦(2007, 2009)的研究中发现, 内隐记忆与外显记忆在编码与提取阶段都存在非对称性, 例如, 编码阶段的干扰影响外显记忆但是对内隐记忆没有影响, 而提取阶段的干扰影响内隐记忆但对外显记忆没有影响。那么这种非对称性也可能存在于胆碱对两种记忆的影响中, 因此在探讨胆碱对两种记忆的影响时, 有必要从记忆加工的不同阶段进行考察。
尼古丁与毒扁豆碱等药物同属于胆碱类药物中的拟胆碱药物, 它与其他拟胆碱药物一样能使人体内胆碱水平上升(抗胆碱药物作用相反)。因此在本研究中, 我们使用拟胆碱药物尼古丁, 检验两种记忆在药物影响下是否产生不同的变化。另外, 有研究者认为, 词干补笔、范例产生等产生式内隐记忆任务更容易混入外显记忆成分(MacLeod, 2008; Sheldon & Moscovitch, 2010), 因此本研究使用词汇判断任务测量内隐记忆, 并使用词汇再认任务测量外显记忆。在实验2中, 分别在编码阶段与提取阶段摄入尼古丁, 观察尼古丁对两种记忆在不同阶段的影响是否不同。通过ERP技术可以观察到与认知加工有关的神经层面的变化, 也由于ERP数据较为敏感, 可以发现行为结果不易观察到的细微变化。因此我们在实验2同时使用ERP技术, 研究记忆过程中受胆碱影响情况的神经机制。
在关于记忆的ERP研究中, N400新旧效应与P600新旧效应常被用作代表内隐记忆与外显记忆的脑电成分。N400新旧效应是在刺激呈现后300~500 ms时间段, 旧刺激引起的平均波幅减去新刺激引起的平均波幅所得到的差异波。当内隐记忆受到影响后, 表现出启动量下降并伴随N400新旧效应消失, 即新旧刺激之间的波幅差异减小。而P600新旧效应为在刺激呈现后500~800ms时间段, 旧刺激引起的平均波幅减去新刺激引起的平均波幅得到的差异波。当外显记忆受到影响后, 将表现出记忆成绩下降并伴随P600新旧效应消失(唐小庭, 2013; 孟迎芳, 郭春彦, 2009)。因此在本研究中, 将重点分析这两个ERP成分是否受到胆碱影响。
2 实验1:尼古丁对内隐记忆与外显记忆的影响
实验1采用词汇判断与词汇再认任务, 比较内隐记忆与外显记忆在有无尼古丁的条件下, 记忆成绩是否有所不同。每位被试都参与尼古丁条件与安慰剂条件, 两种条件分别在两天进行实验。为保证实验结果的稳定性与有效性, 我们在两次实验前都加入与正式实验流程相同的基线实验作为前测, 通过观察前后测与实验条件之间的交互作用, 来分析实验条件对记忆成绩造成的影响。如果尼古丁能对内隐记忆与外显记忆产生影响, 那么尼古丁组的前后测之间的变化趋势可能与安慰剂组的前后测变化趋势不同。
2.1 方法
2.1.1 被试
共32名被试(男性27人, 女性5人), 平均年龄27.1岁。右利手, 视力或矫正视力正常, 身体健康。均自愿参与实验, 实验结束后付与被试一定报酬。2名被试因操作错误而被剔除, 最终30名被试数据进入分析。
2.1.2 材料
低频双字词720个, 选自北京语言学院语言教学研究所编著的《现代汉语频率词典》(1986), 词频为2.3~12.2/百万, 平均词频为3.654/百万。将双字词随机分为24组, 每组30个词。
将24组词随机分到4个测试组中:安慰剂前测、安慰剂后测、尼古丁前测、尼古丁后测, 每个测试组都包含内隐记忆任务与外显记忆任务, 两个记忆任务都各自包含深浅加工, 每个测试组中6组词, 1组为内隐记忆学习阶段的深加工词, 以白色字体颜色呈现; 1组为内隐记忆学习阶段的浅加工词, 一半红色字体呈现, 一半蓝色呈现; 1组作为外显记忆学习阶段的深加工词; 1组作为外显记忆学习阶段的浅加工词; 1组为内隐记忆在测验阶段的新词; 1组为外显记忆在测验阶段的新词。在学习阶段, 内隐记忆与外显记忆深加工词混合呈现, 浅加工词混合呈现。在测验阶段, 外显记忆深、浅加工与新词混合呈现, 内隐记忆深、浅加工与新词混合呈现, 测验阶段字体颜色全部为白色。通过将另外选取的低频双字词前后两个字拆开, 随机组合形成假词, 去除音或义上可能存在的组合, 得到120个假词, 分成4组, 分别在4个实验中的内隐测验阶段与深加工、浅加工、新词混合呈现。所有双字词均为60号黑体。
2.1.3 尼古丁设备
SMOK®电子烟, 电压设定为4 V, halo®烟油, 尼古丁组使用含尼古丁12 mg/ml的烟油, 安慰剂使用含尼古丁0 mg/ml的烟油, 两者在味道与外形上一致。
2.1.4 烟碱依赖量表
本研究使用Fagerstrom尼古丁依赖测试问卷(Fagerstrom Test of Nicotine Dependence Questionnaire, FTND)筛选被试, 问卷共有六个条目。最高分为10分, 最低为0分, 分数越高表示被试尼古丁依赖程度越高。依赖程度根据得分划分五个水平:很低(0~2分), 低(3~4分), 中等(5分), 高(6~7分), 很高(8~10分)。黄晶晶等人对量表在中国地区进行过考究(黄晶晶, 2008), 量表的Cronbach’s α系数达到0.704, 各项目之间相关系数平均0.36, 各项目与总分之间相关系数为0.461~0.819。在效度方面, 量表条目的KMO值达到0.803, 各条目的因子载荷值较高, 量表只提取一个公因子, 可以解释总方差的43.721%, 有较好的结构效度。本研究的被试选取尼古丁依赖程度为低或很低, 即问卷得分0~4分, 平均分1.8。
2.1.5 实验程序
采用Presentation 0.71软件编制实验程序。被试在隔音室内完成个别施测, 显示器背景为黑色, 距离被试80 cm。被试在实验前2小时内不能使用含有尼古丁的物品。
实验分两天进行, 一天进行尼古丁条件, 一天进行安慰剂条件, 每位被试都参与两次实验, 两种条件的顺序在被试间平衡, 被试不知道有一次是安慰剂, 每次实验包括以下11个阶段:
(1)基线安慰剂摄入:使用含尼古丁0 mg/ml的烟油0.1 ml。
(2)学习阶段(前测):分组呈现深加工和浅加工各60个双字词。要求被试对深加工词进行主观愉快度判断, 愉快按F键, 不愉快按J键; 对浅加工词进行颜色判断, 红色按F键, 蓝色按J键。每个词呈现时间为500 ms, 刺激间隔(ISI)为1400~ 1800 ms。
(3)休息阶段:休息10分钟。
(4)内隐测验阶段(前测):30个深加工词、30个浅加工词、30个新词与30个假词混合随机呈现, 要求被试进行真/假词判断, 真词按F键, 假词按J键。当被试按键后词汇消失, 刺激间间隔(ISI)为1400~ 1800 ms。
(5)外显测验阶段(前测):30个深加工词、30个浅加工词与30个新词混合随机呈现, 要求被试进行新/旧词判断, 新词按F键, 旧词按J键。当被试按键后词汇消失, 刺激间间隔(ISI)为1400~ 1800 ms。
(6)尼古丁摄入:在尼古丁条件下, 使用含尼古丁12 mg/ml的烟油0.1 ml; 在安慰剂条件下, 使用含尼古丁0 mg/ml的烟油0.1 ml。
(7)休息阶段:休息10分钟。
(8)学习阶段(后测):流程同前测的学习阶段。
(9)休息阶段:休息10分钟。
(10)内隐测验阶段(后测):流程同前测的内隐测验阶段。
(11)外显测验阶段(后测):流程同前测的外显测验阶段。
实验流程见图1。
图1

2.1.6 数据分析
外显记忆以正确率与反应时为指标, 正确率为旧词的击中率减去新词的虚报率, 反应时为判断正确旧词的平均反应时。内隐记忆以启动量为指标, 启动量为判断正确新词的反应时减去判断正确旧词的反应时。
数据统计分析采用SPSS 17.0软件包进行。
2.1.7 实验伦理
该实验得到福建师范大学伦理委员会同意。
2.2 结果分析
2.2.1 外显记忆结果分析
外显记忆任务中各种条件下的反应时和正确率见表1。
表1 外显记忆任务的正确率和反应时
| 指标 | 深加工编码 | 浅加工编码 | ||||||
|---|---|---|---|---|---|---|---|---|
| 尼古丁 | 安慰剂 | 尼古丁 | 安慰剂 | |||||
| 前测 | 后测 | 前测 | 后测 | 前测 | 后测 | 前测 | 后测 | |
| 正确率 | 0.38(0.21) | 0.23(0.20) | 0.35(0.27) | 0.41(0.25) | 0.16(0.15) | 0.05(0.12) | 0.14(0.11) | 0.09(0.10) |
| 反应时(ms) | 857(149) | 861(147) | 932(206) | 896(201) | 950(238) | 899(275) | 1030(372) | 965(278) |
注:括号内为标准差(下同)
首先我们对正确率进行加工水平(深vs浅) ×实验条件(尼古丁vs安慰剂)×前后测(前测vs后测)的重复测量方差分析。结果发现加工水平主效应显著, F(1,29) = 46.14, p < 0.001, ηp² = 0.614; 实验条件与前后测交互效应显著, F(1,29) = 24.78, p < 0.001, ηp² = 0.461。简单效应分析发现, 在安慰剂条件下前后测之间差异不显著, F(1,29) = 0.03, p = 0.861; 而在尼古丁条件下后测比前测成绩更低, F(1,29) = 32.91, p < 0.001。三者间交互效应显著, F(1,29) = 28.58, p < 0.001, ηp² = 0.496。进一步简单效应分析表明, 在深加工的后测中, 尼古丁条件下的正确率显著低于安慰剂条件, F(1,29) = 22.62, p < 0.001, ηp² = 0.438; 而在前测中两者没有差异, F(1,29) = 0.38, p = 0.542, 说明尼古丁导致深加工条件下, 外显记忆成绩下降。而在浅加工的后测中, 尼古丁条件与安慰剂条件差异不显著, F(1,29) = 2.71, p = 0.110; 同样在前测中两者差异也不显著, F(1,29) = 0.24, p = 0.630, 说明尼古丁对浅加工的影响不大。
随后我们对反应时进行加工水平(深vs浅) ×实验条件(尼古丁vs安慰剂)×前后测(前测vs后测)的重复测量方差分析。结果仅发现加工水平主效应显著, F(1,29) = 6.236, p < 0.05, ηp² = 0.177; 加工水平与前后测交互效应显著, F(1,29) = 4.795, p < 0.05, ηp² = 0.142。而并没有发现三者交互作用, F(1,29) = 0.114, p = 0.738; 也没有发现与实验条件有关的交互效应, 说明尼古丁条件与安慰剂条件之间并没有差异, 表明尼古丁对外显记忆的反应时影响不大。
2.2.2 内隐记忆结果分析
内隐记忆任务中各种条件下的启动量见表2。
表2 内隐记忆任务的启动量
| 指标 | 深加工编码 | 浅加工编码 | ||||||
|---|---|---|---|---|---|---|---|---|
| 尼古丁 | 安慰剂 | 尼古丁 | 安慰剂 | |||||
| 前测 | 后测 | 前测 | 后测 | 前测 | 后测 | 前测 | 后测 | |
| 启动量 | 59(44) | 6(45) | 21(34) | 44(44) | 47(42) | 10(38) | 32(42) | 37(33) |
我们首先对内隐记忆的启动量进行加工水平(深vs浅) ×实验条件(尼古丁vs安慰剂)×前后测(前测vs后测)的重复测量方差分析。结果表明, 实验条件与前后测交互效应显著, F(1,29) = 37.47, p < 0.001, ηp² = 0.564; 在尼古丁条件下, 后测成绩比前测更低, F(1,29) = 19.15, p < 0.001; 而在安慰剂条件下, 后测成绩比前测高, F(1,29) = 5.81, p < 0.05。
三者间交互效应显著, F(1,29) = 6.59, p < 0.05, ηp² = 0.185。进一步简单效应分析表明, 在深加工的尼古丁条件下, 后测的启动量显著小于前测, F(1,29) = 20.33, p < 0.001; 而在安慰剂条件中后测的启动量显著大于前测, F(1,29) = 4.31, p < 0.05, 说明尼古丁导致深加工条件下, 内隐记忆成绩下降。在浅加工的尼古丁条件下, 后测的启动量小于前测, F(1,29) = 17.19, p < 0.001, ηp² = 0.391; 而在安慰剂条件下两者差异不显著, F(1,29) = 0.70, p = 0.409, 说明尼古丁也影响了浅加工条件下的内隐记忆成绩, 但是影响程度没有深加工大。
综上所述, 相对于安慰剂条件, 在尼古丁摄入后, 无论是内隐记忆还是外显记忆, 记忆成绩都有所下降。
3 实验2:尼古丁对记忆不同阶段的影响
实验1发现摄入尼古丁后, 内隐记忆与外显记忆成绩都有一定程度地下降。前人研究已经发现在不同阶段摄入尼古丁对外显记忆的影响不同, 那么对内隐记忆的影响是否也有所不同?编码与提取的非对称性在内隐记忆与外显记忆之间是否一致?为探究这些问题, 实验2分别在编码与提取阶段使用尼古丁, 比较在不同阶段摄入尼古丁对两种记忆影响是否有所不同。在实验2中使用ERP技术, 以进一步了解其中的神经机制与受影响的认知过程。我们推测尼古丁对内隐记忆的不同阶段可能生不同的结果, 并且在ERP结果上, N400新旧效应与P600新旧效应也发生变化。
3.1 方法
3.1.1 被试
共24名被试(男性18人, 女性6人), 平均年龄21.3岁。右利手, 视力或矫正视力正常, 身体健康。均自愿参与实验, 实验结束后付与被试一定报酬。5名被试因叠加数不够而被剔除, 最终19名被试的ERP数据进入分析。
3.1.2 材料
低频双字词1440个, 分为24组, 每组60个, 其余与实验1相同。
3.1.3 尼古丁设备
同实验1。
3.1.4 烟碱依赖量表
同实验1。
3.1.5 实验程序
采用Presentation 0.71软件编制实验程序。被试在隔音室内完成个别施测, 显示器背景为黑色, 距离被试80 cm。被试在实验前2小时内不能使用香烟或其他含有尼古丁的物品。
实验分两天进行, 一次为编码前使用尼古丁, 一次为提取前使用尼古丁, 每位被试都参与两次实验, 两种条件在被试间平衡, 正式实验前同样加入使用过安慰剂的前测, 每次实验包括以下11个阶段:
(1)~(5)步骤同实验1。
(6)尼古丁摄入:在编码条件下, 使用血压计测量血压与心率, 之后使用含尼古丁12 mg/ml的烟油0.1 ml, 过3分钟后再测量血压与心率; 在提取条件下, 被试休息10分钟。
(7)学习阶段:同实验1。
(8)尼古丁摄入:在编码条件下, 被试休息10分钟; 在提取条件下, 被试使用含尼古丁12 mg/ml的烟油0.1 ml, 之后休息, 共10分钟。
(9)内隐测验阶段(实验处理):同实验1。
(10)外显测验阶段(实验处理):同实验1。
实验流程见图2。
图2
3.1.6 行为数据分析
同实验1。
3.1.7 ERP的记录与分析
采用Neuroscan-64导脑电采集分析系统和Ag/AgCl电极帽, 连续记录使用过程中的EEG。电极位置采用10-20扩展电极系统, 单极导联, 以头顶作为参考电极采集数据, 接地点在Fpz和Fz连线的中点, 左眼眶上、下侧2个电极记录垂直眼电, 两眼外侧的2个电极记录水平眼电。滤波带通为0.05~100 Hz, A/D采样率为1000 Hz。脑电采集过程中所有电极与头皮接触电阻均小于5 kΩ。对脑电记录进行脱机迭加平均处理, 分析窗口为-200~ 1200 ms, 用-200~0 ms作为基线进行矫正。伴有眨眼、眼动、肌电等伪迹的数据均被排除, 排除标准为±75 μV。数据离线处理时转化为双侧乳突作 参考。
对于记忆相关的ERP成分, 我们主要分析额区与顶区新旧效应, 新旧效应是指将旧词引发的平均波幅减去新词的平均波幅所得的差异波。额区新旧效应是在额区的300~500 ms时间段(N400)的差异波, 选取F1、Fz、F2三个电极的平均值作为额区的ERP成分; 顶区新旧效应是在顶区的500~ 800 ms时间段(P600)的差异波, 选取P1、Pz、P2三个电极的平均值作为顶区的ERP成分(唐小庭, 2013; 孟迎芳, 郭春彦, 2009)。对每个成分分别进行2(词类型:深、浅)×2(摄入阶段:编码、提取)×2(前后测:前测、后测)的重复测量方差分析。
头皮后部与前部的N100成分受注意显著影响, 因此将F1、Fz、F2电极的平均值作为额区的N100成分进行分析, 将O1、Oz、O2电极的平均值作为枕区的N100成分, 时间段选取50~150 ms, 枕区P250成分主要受早期语义加工影响(赵仑, 2010), 因此将O1、Oz、O2电极的平均值作为枕区的P250成分进行分析, 时间段选取150~300 ms。对每个成分平均波幅分别进行3(词类型:深、浅、新词)×2(摄入阶段:编码、提取)×2(前后测:前测、后测)的重复测量方差分析。
3.2 实验结果
3.2.1 生理指标
在生理指标上, 收缩压在尼古丁摄入前后差异显著, t(23) = -3.20, p < 0.01, Cohen’d = 0.289, 尼古丁摄入后使收缩压上升。舒张压在尼古丁摄入前后差异边缘显著, t(23) = -1.79, p = 0.087, Cohen’d = 0.435, 同样表现为使用尼古丁后血压上升。心率在尼古丁摄入前后差异不显著, t(23) = -1.53, p = 0.140。
3.2.2 行为结果
(1)外显记忆结果分析
外显记忆任务中各种条件下的反应时和正确率见表3。
表3 外显记忆任务的正确率和反应时
| 指标 | 深加工编码 | 浅加工编码 | ||||||
|---|---|---|---|---|---|---|---|---|
| 编码 | 提取 | 编码 | 提取 | |||||
| 前测 | 后测 | 前测 | 后测 | 前测 | 后测 | 前测 | 后测 | |
| 正确率 | 0.33(0.24) | 0.26(0.18) | 0.33(0.18) | 0.28(0.18) | 0.10(0.11) | 0.09(0.09) | 0.11(0.10) | 0.06(0.09) |
| 反应时(ms) | 1015(345) | 814(115) | 925(211) | 860(149) | 1020(347) | 875(150) | 944(184) | 888(191) |
首先我们对正确率进行加工水平(深vs浅)×摄入阶段(编码vs提取)×前后测(前测vs后测)的重复测量方差分析。结果表明, 加工水平主效应显著, F(1,23) = 49.26, p < 0.001, ηp² = 0.682; 前后测主效应显著, F(1,23) = 13.07, p < 0.05, ηp² = 0.362; 后测的记忆成绩低于前测。三者间交互效应边缘显著, F(1,23) = 3.54, p = 0.073, ηp² = 0.133。进一步简单效应分析表明, 在深加工的编码条件下, 后测的正确率显著低于前测, F(1,23) = 6.24, p < 0.05, ηp² = 0.213; 提取条件下前测与后测差异边缘显著, F(1,23) = 3.49, p = 0.075。在浅加工的编码条件下, 前测和后测的差异不显著, F(1,23) = 0.12, p = 0.733; 在提取条件下, 后测的成绩显著低于前测, F(1,23) = 5.52, p < 0.05。
表4 内隐记忆任务的启动量
| 指标 | 深加工编码 | 浅加工编码 | ||||||
|---|---|---|---|---|---|---|---|---|
| 编码 | 提取 | 编码 | 提取 | |||||
| 前测 | 后测 | 前测 | 后测 | 前测 | 后测 | 前测 | 后测 | |
| 启动量 | 26(19) | 18(31) | 24(20) | 1(23) | 21(27) | 28(26) | 19(21) | 3(22) |
随后我们对反应时进行加工水平(深vs浅) × 摄入阶段(编码vs提取) × 前后测(前测vs后测)的重复测量方差分析。结果仅发现加工水平主效应显著, F(1,23) = 6.20, p < 0.05, ηp² = 0.212; 前后测主效应显著, F(1,23) = 6.88, p < 0.05, ηp² = 0.230。其他主效应与交互效应均不显著。
(2)内隐记忆结果分析
内隐记忆任务中各种条件下的启动量见表3。
我们对启动量进行加工水平(深vs浅)×摄入阶段(编码vs提取)×前后测(前测vs后测)的重复测量方差分析。结果表明, 加工水平主效应不显著, F(1,23) = 0.004, p = 0.951; 摄入阶段与前后测交互效应显著, F(1,23) = 7.74, p < 0.05, ηp² = 0.252。进一步简单效应分析表明, 在编码条件下前测与后测差异不显著, F(1,23) = 0.01, p = 0.913; 在提取条件下差异显著, F(1,23) = 16.86, p < 0.001, 说明在提取阶段下, 尼古丁使内隐记忆成绩显著下降。三者交互效应不显著, F(1,23) = 0.29, p = 0.597。但是观察描述性统计可以看出, 在编码条件下, 深加工下前测与后测的趋势与浅加工不同, 因此我们单独对编码条件下的内隐记忆成绩做加工水平(深vs浅) ×前后测(前测vs后测)的重复测量方差分析, 结果发现加工水平与前后测交互效应边缘显著, F(1,23) = 3.14, p = 0.089, ηp² = 0.120, 说明在编码前摄入尼古丁对深浅加工的影响不同。
3.2.3 ERP结果
(1) 外显记忆结果分析
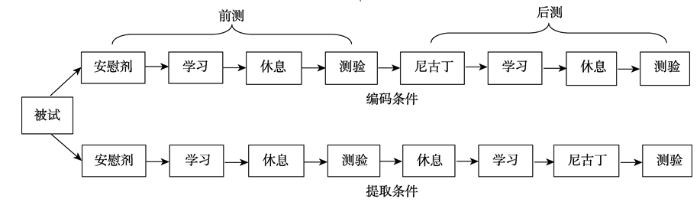
首先我们对记忆相关的ERP成分的差异波进行方差分析, 结果发现在300~500 ms (N400), 摄入阶段与前后测之间交互效应边缘显著, F(1,18) = 4.26, p = 0.054, ηp² = 0.191。进一步简单效应分析结果显示, 在编码条件下, 前测与后测的差异不显著, F(1,18) = 0.57, p = 0.461; 而在提取条件下差异显著, F(1,18) = 4.40, p < 0.05。表现为前测中旧词引起的波幅比新词更正(新旧效应), 而后测中则没有新旧效应, 说明提取前摄入尼古丁N400成分的新旧效应消失。在500~800 ms (P600), 前后测与摄入阶段交互效应边缘显著, F(1,18) = 3.423, p = 0.081, ηp² = 0.160。进行简单效应分析后发现, 在编码条件下, 前测与后测差异不显著, F(1,18) = 0.06, p = 0.809; 在提取条件下, 前测与后测差异显著, F(1,18) = 4.44, p < 0.05。其他主效应与交互效应均不显著。结果见图3。
图3
其次对注意等相关的ERP成分进行分析, 结果发现在额区50~150 ms (N100), 观察到三个变量之间交互作用显著, F(2,17) = 3.82, p < 0.05, ηp² = 0.175。简单效应分析结果发现, 在编码的深加工条件下, 前测与后测的差异显著, F(1,18) = 4.70, p < 0.05; 在提取的深加工条件下差异不显著, F(1,18) = 0.94, p = 0.346。在编码的浅加工条件下差异显著, F(1,18) = 5.60, p < 0.05, ηp² = 0.237; 在提取的浅加工条件下差异不显著, F(1,19) = 2.35, p = 0.143, 说明只有在编码阶段摄入尼古丁, 才影响额区的N100成分, 而提取前摄入则没有影响。而在枕区的50~150 ms (N100), 前后测与摄入阶段的交互作用差异显著, F(1,18) = 4.71, p < 0.05, ηp² = 0.207。进一步简单效应分析结果发现, 在编码条件下前测与后测差异不显著, F(1,18) = 0.05, p = 0.825; 在提取条件下两者差异显著, F(1,18) = 6.29, p < 0.05, ηp² = 0.259, 说明在编码前摄入尼古丁, 对枕区的N100成分没有影响, 而在提取前摄入尼古丁则对枕区N100产生影响。在150~300 ms (P250), 前后测与摄入阶段之间的交互效应显著, F(1,18) = 8.02, p < 0.05, ηp² = 0.308。进一步简单效应分析发现, 在编码条件下, 前测与后测差异不显著, F(1,18) = 2.59, p = 0.125; 在提取条件下, 前测与后测差异边缘显著, F(1,18) = 4.34, p = 0.052, 与枕区N100分成相似, 提取前摄入尼古丁影响P250成分, 而编码前摄入则没有影响。
(2) 内隐记忆结果分析
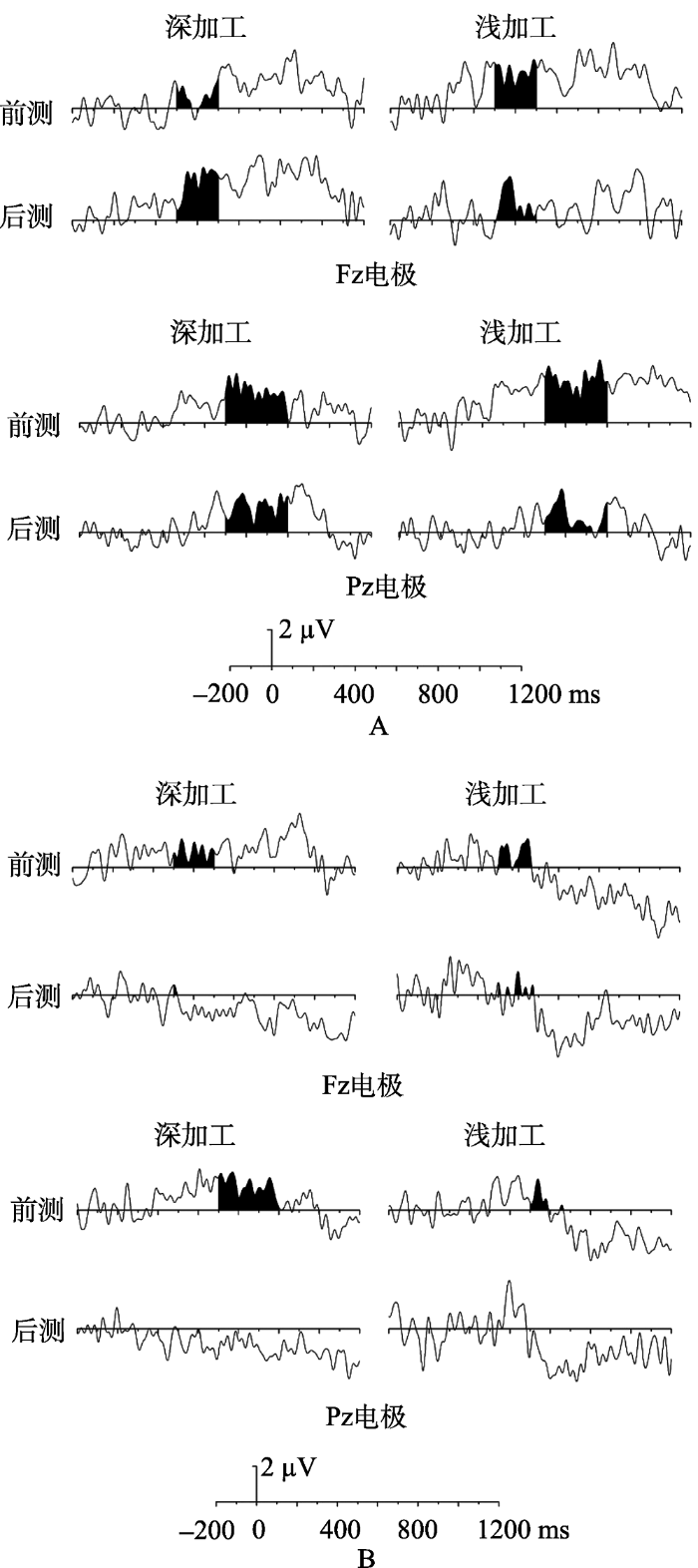
首先, 我们同样对记忆相关的ERP成分的差异波进行方差分析, 结果发现在300~500 ms (N400), 三个变量间交互作用显著, F(1,18) = 5.85, p < 0.05, ηp² = 0.245。进一步简单效应分析结果发现, 在编码前摄入尼古丁的情况下, 深加工条件下的前测与后测之间差异显著, F(1,18) = 4.81, p < 0.05, ηp² = 0.211, 表现为前测中旧词的波幅比新词更正(新旧效应), 而后测中则没有新旧效应。在浅加工条件下, 两者差异不显著, F(1,18) < 0.001, p = 0.996。在提取前摄入尼古丁的情况下, 深加工与浅加工条件下后测中的新旧效应比前测更弱, 但是差异不显著, F(1,18) = 1.32, p = 0.266; F(1,18) = 2.35, p = 0.143 (结果见图4)。
图4
其次再对注意等相关的ERP成分进行方差分析, 在额区的50~150 ms (N100), 各主效应及交互效应均不显著。在枕区的50~150 ms (N100), 仅有摄入阶段主效应边缘显著, F(1,18) = 3.202, p = 0.090, ηp² = 0.151。其他主效应及交互效应均不显著。在150~300 ms (P250), 前后测主效应显著, F(1,18) = 7.35, p < 0.05, ηp² = 0.290。
4 总讨论
本研究采用词汇再认与词汇判断测验, 通过两个实验, 探讨了内隐与外显记忆在尼古丁的影响下, 记忆成绩是否发生变化以及发生怎样的变化。实验1发现, 相对于安慰剂组, 尼古丁的摄入显著降低了随后内隐与外显记忆成绩; 实验2中外显记忆上的结果发现, 尼古丁对深加工下记忆成绩具有更显著的影响。行为结果上, 无论在编码前亦或是提取前摄入尼古丁都降低了深加工条件下的再认成绩; 而浅加工条件的记忆成绩只在提取前摄入尼古丁的情况下受到影响。在ERP结果上, 提取前摄入尼古丁对N400与P600成分造成了与行为类似的削减作用。内隐记忆上的结果发现, 尼古丁同样对深加工下的内隐记忆提取影响更为明显, 行为结果上, 提取前摄入尼古丁对深、浅加工条件的词汇判断成绩构成类似的影响。在ERP结果上, 尼古丁对深加工条件的编码造成了更明显的影响, 使得随后的内隐记忆提取的N400成分产生了相应的降低。
4.1 尼古丁对外显记忆的影响
本研究中发现, 编码前摄入尼古丁使深加工条件下的记忆成绩下降, 这一结果与多数研究不同。前人研究中, 虽然尼古丁同样仅对深加工产生影响, 但是表现为记忆成绩上升, 不同结果的原因可能是由于本研究中摄入胆碱浓度较高导致。Bentley, Driver和Dolan (2011)分析了多项研究发现, 胆碱水平与记忆成绩及相关的大脑激活水平呈倒U型曲线关系。胆碱浓度较低时, 随着浓度升高, 额-顶区的激活程度上升。当超过一定浓度后激活程度反而下降, 相似的情况还发生在视觉皮层与海马。本研究中尼古丁浓度较高(12mg/ml), 因此产生记忆成绩下降的结果。不同于编码条件, 在提取前摄入尼古丁后, 不同加工水平的记忆成绩都有一定程度的下降, 这一点与前人研究一致, 在提取阶段胆碱浓度上升将不利于记忆提取(Rogers & Kesner, 2003; Gais & Born, 2004; Kukolja et al., 2009)。
分析尼古丁对不同加工水平的影响, 本研究的结果也与前人相似(Warburton et al., 2001; FitzGerald et al., 2008), 都表现为深加工条件下记忆成绩受到影响而浅加工不受影响。如前言所述, FitzGerald等人(2008)已通过脑成像研究发现, 尼古丁在编码过程中影响了语义加工的脑区, 而未影响其他非语义加工脑区。另一方面, 在提取前摄入尼古丁时, 尼古丁对不同加工水平的影响没有差异。在编码阶段, 深浅加工采用不同的编码任务, 尼古丁对两种加工水平的记忆成绩产生不同影响。而在提取阶段, 深浅加工条件采用相同的提取任务, 尼古丁则对两种条件都产生影响。可以看出, 尼古丁对记忆的影响与实验任务较为一致。我们推测, 由于两种加工在提取时采用的是相同的任务, 因此调用的是同一模块, 尼古丁影响了提取所调用的模块, 导致记忆成绩发生变化。
观察ERP结果进一步发现, 提取前摄入尼古丁使N400与P600成分都受到影响。最近研究发现N400反映了记忆中的概念启动, 它是指对概念信息的重复加工产生的反应易化, 而P600则与熟悉性有关, 它是当旧刺激再次呈现时对其产生的主观熟悉感(Voss & Paller, 2006; Paller, Voss, & Boehm, 2007; 郭春彦, 高传吉, 李兵兵, 2013)。实验结果中N400成分受到影响, 说明尼古丁影响了外显记忆中自动化的成分。尼古丁不仅影响自动化的概念启动, 也影响了有意识提取的熟悉性(P600)。前人研究发现, 相对于非吸烟者, 吸烟者的前额皮层、右前脑岛等区域在执行任务时激活程度更高, 并伴随任务成绩更差(Sutherland, Ross, Shakleya, Huestis, & Stein, 2011)。Sutherland认为, 这是因为长期吸烟使吸烟者处理信息的效率降低, 需要额外的高级控制操作对任务进行处理, 显示出更强的大脑皮层激活。P600是与外显记忆有关的成分, 同样受到高级控制操作调控, P600成分的波幅受到影响并伴随行为成绩下降, 可能是由于尼古丁降低了被试处理信息时的效率, 导致执行任务时调用了更多的认知资源。
另外, 我们发现在提取前摄入尼古丁条件下, 除了与记忆相关的N400与P600成分受影响外, 与注意相关的N100成分和与早期语义加工相关的P250成分也都受到影响。根据非特异性学说, 胆碱可能通过影响其他心理机能间接地影响记忆。结果说明尼古丁可能不完全是直接对记忆产生影响, 也通过注意等其他心理机能对记忆产生间接的影响。
4.2 内隐记忆
本研究中被试的内隐记忆在尼古丁影响下, 表现出成绩下降的现象, 说明尼古丁同样能对内隐记忆产生影响。本研究的结果与Danion等人(1990)和Schifano和Curran (1994)的结果不一致, 不一致的原因可能由于本研究与前人研究采用的实验范式不同。前文提到同一变量可能对内隐记忆的两种任务产生不同影响, 本研究中采用的词汇判断任务, 与Bentley等人(2003)的实验任务更为相似。进一步分析发现, 在编码前摄入尼古丁对于不同加工水平的内隐记忆成绩的影响不同, 深加工条件下的内隐记忆成绩降低, 浅加工则不受影响, 这一结果与多数关于内隐记忆的理论相违背。以往观点认为, 内隐记忆只与记忆项目的知觉特性相关, 而不涉及概念层面。多重记忆系统理论认为, 内隐记忆依赖于知觉表征系统, 与支持外显记忆行为的情节系统是功能性分离的(Gazzaniga, 1998)。迁移适当加工理论也认为, 内隐记忆主要依赖于知觉加工, 外显记忆主要依赖概念加工(Roediger & McDermott, 1993)。在编码前摄入尼古丁使被试在编码记忆项目时语义加工的脑区受到影响, 进而影响到概念加工过程。而非语义加工脑区不受影响, 因而没有影响知觉加工过程。根据以往观点, 内隐记忆不涉及概念加工仅涉及知觉加工, 以此推论即使概念加工受到影响, 那么内隐记忆成绩也不应受到影响, 但本研究的结果与其不符, 这说明内隐记忆同样也涉及概念加工。另外, 已有研究发现, 呈现与先前学习的项目概念相关的刺激时, 也能引起内隐记忆中的启动效应, 即概念启动, 这也说明内隐记忆不完全依赖于知觉表征。
内隐记忆成绩在提取前摄入尼古丁后消失, 而编码前摄入尼古丁仅仅使深加工条件下的成绩下降, 这说明提取阶段对于尼古丁更为敏感。前言中提到内隐记忆的编码与提取存在非对称性, 编码阶段的干扰对于内隐记忆并不产生影响, 而提取阶段的干扰则使内隐记忆成绩显著下降(孟迎芳, 郭春彦, 2007, 2009; 孟迎芳, 于海莉, 2012), 由此看出内隐记忆在提取阶段更为脆弱, 更易受到其他因素影响。本研究的结果同样也发现内隐记忆在提取阶段的脆弱性, 相对于编码阶段, 提取阶段受尼古丁影响更大。
ERP结果与行为结果一致, 在编码前摄入尼古丁的深加工条件下, 前测与后测在N400成分上差异显著, 并且在行为结果上内隐记忆成绩下降。而在浅加工条件下N400差异不显著, 同时内隐记忆成绩没有受到影响。N400新旧效应是指代内隐记忆的ERP成分, 前人研究发现当内隐记忆受到影响后, 将表现出启动量下降与N400新旧效应消失。另一方面, N400与概念启动有关, 本研究结果中的深加工条件下内隐记忆成绩下降, 并伴随N400成分受到影响。印证了编码前摄入尼古丁损害了编码过程中的概念加工过程, 使随后内隐记忆提取受到影响。
在提取前摄入尼古丁条件下, 后测的N400新旧效应比前测低, 但是差异不显著, 而内隐记忆成绩下降程度却远高于编码阶段。说明尼古丁对内隐记忆提取阶段的影响, 并不仅仅是对记忆的提取过程产生直接的影响。在P250成分上可以观察到前后测的主效应, 表现为后测中P250成分的波幅比前测更低, 说明尼古丁影响了提取过程中的早期语义加工。由于内隐记忆的提取阶段相对于编码阶段
更易受到干扰, 因此提取前摄入尼古丁条件下内隐记忆成绩下降程度更大可能是由于语义加工受到影响导致。
4.3 内隐记忆与外显记忆的对比
对比内隐与外显记忆的结果可以发现, 两者在尼古丁的影响下表现出一定的相似性, 但是也略有不同。编码前摄入尼古丁使两种记忆在深加工条件下的记忆成绩下降, 但是对浅加工没有影响。提取前摄入尼古丁使两种记忆在两种加工水平上都下降, 而影响程度不同。对于本研究的结果, 使用Moscovitch等人提出的成分-加工模型(component of processing model)可以较好地解释。该理论认为记忆过程受三个重要的系统调节:新皮层系统, 内侧颞叶/海马系统和前额皮层/中央系统。其中, 新皮层系统包含知觉和语义模块, 无论内隐记忆还是外显记忆, 在编码阶段都依赖该系统对刺激事件进行登记并转化成前语义表征, 之后进入中央系统进行加工, 最后进入海马系统储存信息。内隐记忆中的重复启动效应主要依赖于该系统在提取阶段对过去信息的再激活。而外显记忆同时受内侧颞叶/海马系统和前额皮层/中央系统的调节。外显提取经由海马系统信息的调出, 随后进入中央系统加工并提取。内隐记忆与外显记忆在编码阶段的任务相同, 因此编码所使用的模块相同, 若尼古丁影响语义模块的登记, 那么在深加工条件下无论是内隐记忆还是外显记忆都会被影响。而知觉模块没有受到影响, 因此浅加工条件下两种记忆成绩都不会受到影响。两种记忆在提取阶段由于任务不同调用不同的模块, 而尼古丁对不同的提取模块影响不同, 因此发生两种记忆之间的分离现象。但是在同一种记忆中, 两种加工方式在提取时调用相同的模块, 因此在加工水平上没有发生分离。
关于内隐记忆与外显记忆的分离现象, 不同观点主要分歧为两者是否从属于不同记忆系统。若两者属于不同的记忆系统, 那么在尼古丁的影响下两种记忆将会发生明显不同的变化, 但是本研究中编码前摄入尼古丁对两种记忆的影响较为一致, 仅在提取前摄入尼古丁条件下略有不同, 因此并不符合该观点。相反, 本研究结果中尼古丁对记忆影响的结果与任务类型具有较高一致性, 即尼古丁对记忆的影响随任务类型的不同发生变化, 结果更符合两者属于同一记忆系统的观点。因此内隐记忆与外显记忆的内在生理机制可能有较大程度的重叠。
参考文献
Effects of handedness and depth of processing on the explicit and implicit memory
DOI:10.1016/j.sbspro.2012.01.005
URL
[本文引用: 2]

In this research the effects of handedness and depth of processing on the explicit and implicit memory was evaluated. An experimental research was carried out individually on 80 students using computerized measurement tools and Edinburgh Handedness Inventory. The data were analyzed through Variance Analysis Method. Results showed that the difference between examinees under different depths of processing is significant in explicit memory. Moreover, there was also a significant difference in explicit memory between the three groups. Mixed-handed and left-handed participants had better recall performance but there was no significant difference in implicit memory. Better performance of the mixed-handed in explicit memory recall test indicates that explicit memory function is dependent on the interrelation between the two hemispheres.
Modulation of fusiform cortex activity by cholinesterase inhibition predicts effects on subsequent memory
DOI:10.1093/brain/awp176 URL [本文引用: 2]
Cholinergic modulation of cognition: Insights from human pharmacological functional neuroimaging
DOI:10.1016/j.pneurobio.2011.06.002 URL [本文引用: 1]
Cholinergic enhancement modulates neural correlates of selective attention and emotional processing
DOI:10.1016/S1053-8119(03)00302-1 URL [本文引用: 3]
Implicit versus explicit retrieval of surnames of famous people: Dissociative effects of levels of processing and age
Long-lasting cognitive improvement with nicotinic receptor agonists: Mechanisms of pharmacokinetic-pharmacodynamic discordance
DOI:10.1016/j.tips.2005.05.007 URL [本文引用: 1]
Memorysystems, processing modes, and components: Functional neuroimaging evidence
DOI:10.1177/1745691612469033 URL [本文引用: 1]
Effects of scopolamine, trimipramine and diazepam on explicit memory and repetition priming in healthy volunteers
DOI:10.1007/BF02244116 URL [本文引用: 5]
Cholinergic modulation of hippocampal activity during episodic memory encoding in postmenopausal women: A pilot study
DOI:10.1097/gme.0b013e3181e04db9 URL [本文引用: 2]
Effects of donepezil on verbal memory after semantic processing in healthy older adults
Low acetylcholine during slow- wave sleep is critical for declarative memory consolidation
DOI:10.1073/pnas.0305404101
URL
PMID:14766981
[本文引用: 3]
The neurotransmitter acetylcholine is considered essential for proper functioning of the hippocampus-dependent declarative memory system, and it represents a major neuropharmacological target for the treatment of memory deficits, such as those in Alzheimer's disease. During slow-wave sleep (SWS), however, declarative memory consolidation is particularly strong, while acetylcholine levels in the hippocampus drop to a minimum. Observations in rats led to the hypothesis that the low cholinergic tone during SWS is necessary for the replay of new memories in the hippocampus and their long-term storage in neocortical networks. However, this low tone should not affect nondeclarative memory systems. In this study, increasing central nervous cholinergic activation during SWS-rich sleep by posttrial infusion of 0.75 mg of the cholinesterase inhibitor physostigmine completely blocked SWS-related consolidation of declarative memories for word pairs in human subjects. The treatment did not interfere with consolidation of a nondeclarative mirror tracing task. Also, physostigmine did not alter memory consolidation during waking, when the endogenous central nervous cholinergic tone is maximal. These findings are in line with predictions that a low cholinergic tone during SWS is essential for declarative memory consolidation.
Brain and conscious experience
FN400 effect: Conceptual processing in explicit memory test
DOI:10.3724/SP.J.1042.2013.01521
URL
Conceptual priming and familiarity are hot spots of researches on neuro-mechanisms of implicit and explicit memory.Supporters of dual-process theories put forward that brain potentials at pre-frontal areas between 300 and 500ms(FN400) are associated with familiarity.However,supporters of singleprocess theories think that studies supporting FN400 as index of familiarity did not take into account the contamination of conceptual priming on explicit memory.Through a series of studies,Paller and his colleagues proved that FN400 is associated with conceptual priming but not familiarity and that familiarity is indexed by brain potentials at parietal areas between 500 and 800ms(LPC).So far,there are still debates on the nero-mechanism of familiarity and conceptual priming.Further researches should explore new methods to dissociate familiarity and conceptual priming.The neuro-mechanism of effects of fluency on familiarity should also be investigated.
Fn400效应: 外显记忆测量中的概念启动加工
DOI:10.3724/SP.J.1042.2013.01521
URL
[本文引用: 1]
Conceptual priming and familiarity are hot spots of researches on neuro-mechanisms of implicit and explicit memory. Supporters of dual-process theories put forward that brain potentials at pre-frontal areas between 300 and 500ms (FN400) are associated with familiarity. However, supporters of single- process theories think that studies supporting FN400 as index of familiarity did not take into account the contamination of conceptual priming on explicit memory. Through a series of studies, Paller and his colleagues proved that FN400 is associated with conceptual priming but not familiarity and that familiarity is indexed by brain potentials at parietal areas between 500 and 800ms (LPC). So far, there are still debates on the nero-mechanism of familiarity and conceptual priming. Further researches should explore new methods to dissociate familiarity and conceptual priming. The neuro-mechanism of effects of fluency on familiarity should also be investigated.
The study on the addiction of smoking behavior in urban residents (Unpublished master’s thesis)
Unaware learning versus preserved learning in pharmacologic amnesia: Similarities and differences
Fmri evidence for dissociation between priming and conscious recognition
DOI:10.1142/S0219635214500149
URL
PMID:25164357
[本文引用: 1]
Evidence from recent studies challenge traditional memory system accounts of separate mechanisms for implicit and explicit memory. The motive behind the present study was to further investigate whether common activations can be detected during explicit and implicit memory performance when using the same procedure for both tasks. Functional magnetic resonance imaging (fMRI) showed separate neural activation to perceptual repetition priming and perceptual recognition of real and nonsense objects, both in terms of the brain regions involved and in the direction of repeated-novel activations. Repetition priming showed deactivation for repeated stimuli in regions not overlapping regions activated in conscious recognition, activation patterns in the two tasks involved largely separate networks. Common activations were observed in three areas, considered as being involved in processes such as stimulus analyses, task monitoring and attention, i.e., processes external to memory. We interpret this as indicating an ...
Cholinergic stimulation enhances neural activity associated with encoding but reduces neural activity associated with retrieval in humans
DOI:10.1523/JNEUROSCI.0203-09.2009 URL [本文引用: 3]
Nicotinic effects on cognitive function: Behavioral characterization, pharmacological specification, and anatomic localization
DOI:10.1007/s00213-005-0164-7 URL [本文引用: 1]
Effects of interference on retrieval process in implicit memory
DOI:10.3724/SP.J.1041.2017.00897
URL
A wide range of studies have shown that executing the other secondary task during encoding has little influence on implicit memory(repetition priming). Somewhat surprisingly, relatively few studies discussed the effects of interference on implicit memory during retrieval, which was confirmed as a process distinct from encoding, but also important in memory. Furthermore, an open question remains as to whether the retrieval interference could affect implicit memory? The effect of interference during retrieval was assessed by comparing a divide-attention(DA) condition, on which participants carried out a memory task(lexical decision) and an interference task(even-odd decision) simultaneously or successively, with a full-attention(FA) condition, on which participants performed only the memory task or interference task. Each experiment consisted of five parts: a study phase, a distraction phase, twice FA interference phases, and a retrieval phase, which included above two types attention conditions. Thirty-five undergraduate students participated in Experiment 1, which investigated whether synchronous interference task during retrieval could affect implicit memory. Experiment 2 further examined whether asynchronous interference could also change the priming of implicit retrieval. Thirty participants took part in Experiment 3, which was designed to examine whether such interference could affect explicit memory retrieval. Therefore, it replaced the lexical decision with recognition task based on Experiment 1. Participants were instructed to make response to corresponding items by pressing keyboard, and were told that the memory and interference task were equally important. They were asked to perform both tasks as quickly and accurately as possible. The Reaction Time(RT) and Accuracy data in retrieval phase were recorded, in order to assess priming effects, the effect of interference and interference task costs. The results showed that, first, the repetition priming results(the facilitation or bias in processing of studied items) were quite consistent across Experiment 1 and 2 both on RT and Accuracy. It reflected that priming would be impacted by interference task, whatever the distraction and memory stimulus presented synchronously or asynchronously. Second, In Experiment 3, there was non-significant difference across attention conditions in recognition Accuracy. Third, we followed Lozito and Mulligan's(2010) method for examining interference task costs. They proposed two measures to obtain distracting task costs for verifying the effects of interference. One of them would work out global costs, which was assessed by comparing performance on the interference task when performed under DA to FA. We found global costs occurred among three experiments, indicating that attention resource competition happened across dual tasks. But significant specific costs, which comparing performance on interference among DA, was only found in Experiment 3, indicating that explicit retrieval would break secondary task performance, whereas, implicit retrieval seemingly has little impact on interference task, but easily influenced by interference task. In conclusion, results from the current study revealed that implicit memory priming could not be regarded as an automatic form of retrieval with ease. And it's necessary for memory retrieval to catch enough cognitive resources. If limited resource was occupied by the other task, implicit retrieval processing would be impacted.
提取干扰对内隐记忆的影响
DOI:10.3724/SP.J.1041.2017.00897
URL
[本文引用: 2]
近三十年来,内隐记忆是认知心理学领域倍受关注的研究方向,吸引了很多专家学者的目光。内隐记忆指的是对先前获得信息的无意识提取,它与对过去经验有意识、有目的回忆的外显记忆是相对应的。关于内隐记忆的影响因素有很多,其中,干扰因素是最为重要的一个因素。当前,很多研究集中于探讨编码干扰对内隐记忆的影响,也得出了较为一致的结论,即编码干扰会影响内隐记忆。然而,关于内隐记忆提取干扰效应的研究很少,而且结论也不一致。 为了进一步探究提取干扰在内隐记忆中所扮演的角色,我们设置了两种不同的干扰任务:知觉干扰、概念干扰,并引入了加工水平这一变量,通过行为实验及事件相关电位(ERP)实验进行研究。行为实验结果表明,在无干扰条件下,词汇判断任务存在着明显的启动效应;而在干扰条件下,这种启动效应受到了影响,知觉干扰条件下深加工启动效应消失,但仍存在浅加工启动效应,概念干扰条件下不存在任何效应。脑电行为结果与行为实验结果一致。ERPs结果表明,无干扰条件下虽没发现浅加工启动效应的ERP成分,然而在深加工启动中,却有明显代表内隐记忆的N400效应;而在干扰条件下,新旧词间的ERPs却没有显著差异。 综合行为与ERP研究结果,我们认为内隐记忆受到了提取干扰的影响,这种影响在不同干扰任务类型中有所差异,概念干扰任务的破坏程度大于知觉干扰任务。
Exploring the role of attention during implicit memory retrieval
DOI:10.1016/j.jml.2010.06.007
URL
[本文引用: 1]
The role of attention during implicit memory retrieval was assessed using a test-phase division of attention. Implicit retrieval is dissociable into perceptual and conceptual forms. Implicit retrieval is further dissociable into tests that involve stimulus identification or stimulus production. The present study used implicit tests that varied on these dimensions. Experiment 1 used a perceptual identification test; Experiment 2 used a word-stem completion test; and Experiment 3 used a category-exemplar production test. Attention was divided with one of several secondary tasks. None of the secondary tasks reduced levels of priming for any of the implicit tests. Furthermore, implicit retrieval generally facilitated secondary task performance rather than producing secondary task costs as is typical with explicit memory. A fourth experiment indicated that explicit recall was negatively affected by most of the secondary tasks, and exhibited a different pattern of secondary task costs. All of the above is consistent with the idea that implicit retrieval is automatic.
The asymmetric effect of interference at encoding or retrieval on implicit and explicit memory
编码与提取干扰对内隐和外显记忆的非对称性影响
以往研究表明在外显记忆中,编码与提取加工存在着非对称性,但在内隐记忆中,二者的关系并不明确,因此该实验采用“学习-再认”范式,考察在编码或提取中分别附加的干扰任务对词汇判断或再认产生的影响。结果证实编码与提取干扰对内隐或外显记忆都具有非对称性的影响,但又存在着差异,即编码干扰会导致随后外显记忆成绩显著减少,而提取干扰对其影响较小,相反,编码干扰对随后内隐测验中启动效应的影响较小,但提取干扰会破坏启动效应,从而为内隐记忆和外显记忆的分离提供了进一步的证据。
The asymmetric relationship between encoding and retrieval in implicit and explicit memory
DOI:10.3724/SP.J.1041.2009.00694 URL [本文引用: 3]
内隐与外显记忆的编码与提取非对称性关系
Encoding and retrieval are two important phases of memory. Encoding produces memory engraved and re-trieval reactivates previously encoded information. The approach to the relationship between encoding and re-trieval is meaningful for the better understanding of the nature of memory. As for explicit memory, the relation of encoding and retrieval has been investigated by many researchers who have found out that the performance of a secondary task during encoding reduced the later memory performance, but the division of the attention in the same way during retrieval had virtually no effect on memory performance, which indicated an asymmetry be-tween encoding and retrieval processes. With respect to implicit memory, an asymmetry was also found in one of our previous studies (Meng & Guo, 2006), but the asymmetry is somewhat different from that in explicit mem-ory, that is, the performance of a concurrent task during encoding had no effect on later task performance, but interference during retrieval disrupted priming, and reduced task performance. In order to clarify the issues we conducted an ERP experiment. The experiment was conducted with 16 undergraduate students as subjects and Chinese characters as stim-uli. The experiment adopted a study-to-test paradigm, in which participants performed a “shallow” (color) study task or a “deep” (pleasant) study task, followed by either a lexical decision (implicit) test (section 1) or a recog-nition (explicit) test (section 2). In interference task participants were asked to account the total number of “+” in a regulated orientation which appeared with a word, and was performed concurrently with either the encoding or the retrieval phase of the memory task for encoding interference condition or retrieval interference condition. The results showed that: (1) the effects of interference to encoding in implicit memory test were different from that to retrieval. Interference during encoding had virtually no effect on N400 or P600 old/new ERP components, but interference during retrieval significantly reduced N400 old/new and P600 old/new components, which ab-olished behavior priming. (2) The effects of interference to encoding on ERP components in explicit memory test were different from that to retrieval. Interference during encoding had significant influence on later P600 old/new effect, but interference during retrieval did not produce such effect. The results confirmed the finding of Meng & Guo (2006), that is, interference during encoding had effect on explicit memory, but left implicit memory intact. Interference during retrieval affected implicit memory, but had little effect on explicit memory. So the relationship between encoding and retrieval was different between implicit and explicit memory, thus providing further evidence on the dissociation between implicit and explicit memory.
The dissocciation between encoding and retrieval in implicit and explicit memory
Encoding and retrieval are two important phases of memory.The study investigated the effects of encoding-interference or retrieval-interference on beheavior in implicit memory test and explicit memory test.The results showed that:(1) the effects of interference to encoding on priming in implicit memory test were different from that to retrieval.Compared with encoding-interference,retrieval-interference had more effect on priming,which indicated an asymmetry between encoding and retrieval processes,and the asymmetry was the same between shallow and deep study tasks.(2) The effects of interference to encoding on old/new effect in explicit memory test were different from that to retrieval.Compared with retrieval-interference,encoding-interference had more effect on old/new effect,which indicated an asymmetry between encoding and retrieval processes,and the asymmetry was more distinct on deep study task.The results supported the asymmetric relationship between encoding and retrieval in implicit and explicit memory,and the asymmetric relationship is different between these two Memory types,which indicates the dissociation between encoding and retrieval and the dissociation between implicit and explicit memories.
Emotion effects on implicit and explicit musical memory in normal aging
DOI:10.1037/pag0000116
URL
PMID:27599018
[本文引用: 1]
Abstract Normal aging affects explicit memory while leaving implicit memory relatively spared. Normal aging also modifies how emotions are processed and experienced, with increasing evidence that older adults (OAs) focus more on positive information than younger adults (YAs). The aim of the present study was to investigate how age-related changes in emotion processing influence explicit and implicit memory. We used emotional melodies that differed in terms of valence (positive or negative) and arousal (high or low). Implicit memory was assessed with a preference task exploiting exposure effects, and explicit memory with a recognition task. Results indicated that effects of valence and arousal interacted to modulate both implicit and explicit memory in YAs. In OAs, recognition was poorer than in YAs; however, recognition of positive and high-arousal (happy) studied melodies was comparable. Insofar as socioemotional selectivity theory (SST) predicts a preservation of the recognition of positive information, our findings are not fully consistent with the extension of this theory to positive melodies since recognition of low-arousal (peaceful) studied melodies was poorer in OAs. In the preference task, YAs showed stronger exposure effects than OAs, suggesting an age-related decline of implicit memory. This impairment is smaller than the one observed for explicit memory (recognition), extending to the musical domain the dissociation between explicit memory decline and implicit memory relative preservation in aging. Finally, the disproportionate preference for positive material seen in OAs did not translate into stronger exposure effects for positive material suggesting no age-related emotional bias in implicit memory. (PsycINFO Database Record (c) 2016 APA, all rights reserved).
Validating neural correlates of familiarity
DOI:10.1016/j.tics.2007.04.002
URL
PMID:17475539
[本文引用: 1]
Familiarity is a pervasive memory phenomenon that occurs in its most basic form when someone recognizes a repeated stimulus without recollecting other aspects of the requisite prior learning episode. Theoretical controversy currently abounds with respect to both the cognitive and neural characteristics of familiarity. Here, we show that the extant data, particularly brain-potential data, are insufficient for validating putative neural correlates of familiarity, and we outline strategies for making progress on this problem. Conceptual priming is an implicit-memory phenomenon that often occurs together with familiarity; experiments that conflate the two phenomena can be misleading. Avoiding this conflation is required to understand familiarity and to determine the extent to which the neurocognitive processes that support priming also drive familiarity.
Effects of divided attention at retrieval on conceptual implicit memory
DOI:10.3389/fpsyg.2016.00005
URL
PMID:26834678
[本文引用: 1]
This study investigated whether conceptual implicit memory is sensitive to process-specific interference at the time of retrieval. Participants performed the implicit memory test of category exemplar generation (Experiments 1 and 3), or the matched explicit memory test of category-cued recall (Experiment 2), both of which are conceptually-driven memory tasks, under one of two divided attention (DA) conditions in which participants simultaneously performed a distracting task. The distracting task was either syllable judgments (dissimilar processes), or semantic judgments (similar processes) on unrelated words. Compared to full attention (FA) in which no distracting task was performed, DA had no effect on category exemplar generation priming overall, but reduced category-cued recall similarly regardless of distractor task. Analyses of distractor task performance also revealed differences between implicit and explicit memory retrieval. The evidence suggests that, whereas explicit memory retrieval requires attentional resources and is disrupted by semantic and phonological distracting tasks, conceptual implicit memory is automatic and unaffected even when distractor and memory tasks involve similar processes.
Cholinergic modulation of the hippocampus during encoding and retrieval
DOI:10.1016/S1074-7427(03)00063-7
URL
PMID:14521875
[本文引用: 4]
The present experiments were aimed at determining whether acetylcholine (ACh) plays a role in encoding and retrieval of spatial information using a modified Hebb illiams maze. In addition, the present experiments tested two computational models of hippocampal function during encoding and retrieval using a maze sensitive to hippocampal disruption. Thirty male, Long-Evans rats served as subjects. Chronic cannulae were implanted bilaterally into the CA3 ( n=26) and CA1 ( n=5) subregions of the hippocampus. Rats were tested using a modified Hebb-Williams maze. In the first experiment, rats were injected with either saline or scopolamine hydrobromide 10 min before testing for each day. The number of errors made per day per group was used as the measure of learning. Encoding was assessed by the average number of errors made on the first five trials of Day 1 compared to the last five trials of Day 1, whereas the average number of errors made on the first five trials of Day 2 compared to the last five trials of Day I was used to assess retrieval. No deficit was found for the saline group. The scopolamine group showed a deficit in encoding, but not retrieval. In the second experiment, rats were injected with either saline or physostigmine 10 min before testing each day. In contrast to the scopolamine groups, the physostigmine group showed a deficit in retrieval, but not encoding. To test whether the retrieval deficit was due to a disruption in storage or gaining access to the information two groups of rats received either saline on Day 1 and physostigmine on Day 2 or physostigmine on Day 1 and saline on Day 2. In addition, one group received physostigmine immediately after testing on Day 1. Data indicate that physostigmine causes a disruption of retrieval by means of a disruption in consolidation process. In conclusion, the cholinergic antagonist, scopolamine, disrupts encoding in both CA3 and CA1 subregions of the hippocampus. Furthermore, the cholinesterase inhibitor, physostigmine, boosts ACh action during a time when cholinergic levels need to decline for proper consolidation.
Regional brain activity during shape recognition impaired by a scopolamine challenge to encoding
DOI:10.1046/j.1460-9568.1999.00784.x
URL
PMID:10564377
[本文引用: 1]
Abstract In the present positron emission tomography (PET) study, we examine the effect of a scopolamine-induced challenge to encoding upon the pattern of regional cerebral blood flow during recognition of a list of abstract visual shapes 3 days after encoding of these shapes. This study was conducted to test hypotheses concerning the fusiform and thalamic contributions to object recognition arising from a previous imaging study of impaired recognition. In that study, we demonstrated that activity in the fusiform cortex and the thalamus during shape recognition was modulated by memory challenges. These memory challenges included, on one hand, impaired storage as a consequence of diazepam administration during encoding, and, on the other hand, impaired retrieval caused by a perceptual challenge. Activation in the fusiform cortex decreased during impaired recognition, irrespective of the type of challenge. In contrast, thalamic activation increased only when the recognition deficit resulted from impaired memory storage. Based on these results, we hypothesized that fusiform activation during recognition reflects the matching of an incoming stimulus with a stored one, whereas thalamic activation reflects retrieval attempts. These hypotheses would receive considerable support if scopolamine, which also impairs memory storage, induced similar modulations of fusiform and thalamic activation. In the present study, we observed that a scopolamine challenge to encoding does indeed modulate the activity in the very same regions that were previously modulated by a diazepam challenge. Hence, a similar memory deficit, although primarily effected through different neurochemical pathways, was paralleled by a similar modulation of activity in the same set of nodes in the shape recognition network. In the fusiform cortex, scopolamine decreased recognition-related activity, as did the sensory challenge of retrieval. Furthermore, covariate analysis demonstrated that the level of fusiform activity linearly correlates with behavioural performance. In the thalamus, activation increased following impaired encoding. This is in accordance with the idea that enhanced thalamic activity reflects increased effort expended in retrieval. In addition, in the intraparietal sulcus, differential activation also increased following impaired memory storage, possibly reflecting enhanced visuospatial attention in an effort to compensate for impaired performance.
Dissociation of the neural correlates of implicit and explicit memory
DOI:10.1038/33396 URL [本文引用: 1]
Pharmacological models of memory dysfunction? A comparison of the effects of scopolamine and lorazepam on word valence ratings, priming and recall
DOI:10.1007/BF02245086 URL [本文引用: 4]
Recollective performance advantages for implicit memory tasks
DOI:10.1080/09658211.2010.499876
URL
PMID:20721805
[本文引用: 1]
A commonly held assumption is that processes underlying explicit and implicit memory are distinct. Recent evidence, however, suggests that they may interact more than previously believed. Using the remember now procedure the current study examines the relation between recollection, a process thought to be exclusive to explicit memory, and performance on two implicit memory tasks, lexical decision and word stem completion. We found that, for both implicit tasks, words that were recollected were associated with greater priming effects than were words given a subsequent familiarity rating or words that had been studied but were not recognised (misses). Broadly, our results suggest that non-voluntary processes underlying explicit memory also benefit priming, a measure of implicit memory. More specifically, given that this benefit was due to a particular aspect of explicit memory (recollection), these results are consistent with some strength models of memory and with Moscovitch's (2008) proposal that recollection is a two-stage process, one rapid and unconscious and the other more effortful and conscious.
Chronic smoking, but not acute nicotine administration, modulates neural correlates of working memory
DOI:10.1007/s00213-010-2013-6 URL [本文引用: 1]
The effect of interferenceatretrieval on implicit memory (Unpublished master’s thesis)
提取干扰对内隐记忆的影响
(硕士学位论文).
Age and time effects on implicit and explicit learning
DOI:10.1080/0361073X.2014.926778
URL
PMID:25054644
[本文引用: 1]
Background/Study Context: It has been proposed that effects of aging are more pronounced for explicit than for implicit motor learning. The authors evaluated this claim by comparing the efficacy of explicit and implicit learning of a movement sequence in young and older adults, and by testing the resilience against fatigue and secondary tasking after learning. It was also examined whether explicit learning in older adults can be promoted by alleviating time constraints during learning. Methods: The alternating serial reaction time task (ASRTT) was used. Experiment 1 compared the benefits of receiving full instructions about the stimulus sequence relative to receiving no instructions in young (2025 years) and older (5065 years) adults during retention and during transfer to fatigue and secondary task conditions. Experiment 2 alleviated time constraints during the initial bouts of practice with full instructions. Results: Experiment 1 indicated that the older adults learned on the ASRTT and achieved similar performance as young adults when no instructions were given. In contrast to the young adults, learning was not superior in older adults who received full instructions compared with those who did not. Experiment 2 indicated that alleviating time constraints allowed some of the older adults to gain from instruction but only under relatively low time constraints, but there was no retention with rigorous time constraints. Conclusion: Explicit learning, but not implicit learning, declines in older adults. This is partly due to older adults difficulties to apply explicit knowledge. Less rigorous time constraints can help to ameliorate some of these difficulties and may induce levels of explicit learning in older adults that will result in superior performance compared with implicit learning. Implicit learning did occur under time constraints that prevented explicit learning.
Fluent conceptual processing and explicit memory for faces are electrophysiologically distinct
DOI:10.1523/JNEUROSCI.3931-05.2006
URL
PMID:16421312
[本文引用: 2]
Abstract Implicit memory and explicit memory are fundamentally different manifestations of memory storage in the brain. Yet, conceptual fluency driven by previous experience could theoretically be responsible for both conceptual implicit memory and aspects of explicit memory. For example, contemplating the meaning of a word might serve to speed subsequent processing of that word and also make it seem familiar. We examined electrophysiological correlates of conceptual priming with 180 celebrity faces to determine whether or not they resemble electrophysiological correlates of explicit memory. Celebrity faces are ideal for this purpose because they carry with them preexisting conceptual information (i.e., biographical facts) that can selectively be brought to mind such that conceptual processing can be manipulated systematically. In our experiment, exposure to biographical information associated with only one-half of the celebrities yielded conceptual priming for those faces, whereas all faces were perceptually primed. Conceptual priming was indexed by positive brain potentials over frontal regions from approximately 250 to 500 ms. Explicit memory retrieval was associated with later brain potentials over posterior regions that were strikingly similar to potentials previously associated with pure familiarity for faces (when a face seems familiar in the absence of retrieval of any specific information about previous occurrence). Furthermore, the magnitude of conceptual priming was correlated across subjects with the amplitude of frontal but not posterior potentials, whereas the opposite was true for explicit memory. Distinct brain processes were thus associated with conceptual priming and conscious recognition of faces, thus providing a sharper focus on the border between implicit and explicit memory.
Improved incidental memory with nicotine after semantic processing, but not after phonological processing
DOI:10.1007/s002130000565 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}