

1 引言
在注意系统中存在一种现象, 对曾经注意过的位置或物体会发生一个先易化后抑制的过程, 这个抑制的过程被称为返回抑制(inhibition of return, IOR) (Posner & Cohen, 1984)。该过程可以帮助人们避免关注曾经注意过的物体或位置, 从而偏向新颖的物体或位置来提高搜索效率, 减少不必要的搜索过程(Klein, 2000; Redden et al., 2021; Wang et al., 2010)。关于IOR的产生阶段问题, 一直存在两种观点, 一种是知觉抑制理论, 这一理论认为IOR现象阻碍对曾经搜索过的位置进行再次注意, 使注意重新定向到其他未搜索过的位置, 这种现象是对注意的本身进行的一种抑制, 即IOR是一种注意后效, 能够影响刺激的早期知觉加工。已有研究发现, 反映在早期知觉阶段受到加工的P1和N1成分的波幅在无效线索化条件下增强, 验证了IOR是通过抑制对有效线索位置刺激的早期知觉加工而产生的观点(Hopfinger & Mangun, 2001; Prime & Jolicœur, 2009)。另一种反应抑制理论则认为, IOR仅是一种避免对线索化位置做出反应的运动偏向, 表现为延迟对线索化位置的目标进行反应。以往研究结果也发现在有效线索化条件下代表反应抑制成分(Go/Nogo-N2)产生的波幅小于无效线索化条件, 且出现的时间更早(Taylor & Klein, 1998; Tian & Yao, 2008)。
注意系统中的IOR现象可以有效的帮助人们偏向新颖刺激, 提高搜索效率(Klein, 2000; Lupiáñez et al., 2006)。此外, 已有研究发现人类对情绪刺激具有高度敏感性, 它能够引起人类的注意偏向进而获得优先加工, 尤其是与人类生存密切相关的恐惧等负性情绪刺激(Anderson & Phelps, 2001; Pourtois et al., 2013)。既然IOR与情绪刺激均能够引起注意偏向, 提高搜索效率, 这两者之间是否可以存在相互影响, 引起了研究者的关注。情绪与IOR相关研究的焦点集中在IOR经典范式下产生的IOR效应是否受到情绪刺激的影响, 但以往研究的结果并不一致(Berdica et al., 2017; Jia et al., 2019; Pérez-Dueñas et al., 2009, 2014; Rutherford & Raymond, 2010; Silvert & Funes, 2016)。考察二者之间关系的研究, 大致分为两类:一类研究将情绪刺激呈现在IOR经典范式中的线索位置, 少数研究结果发现负性线索后的IOR效应量减少(Okon-Singer et al., 2020; Pan et al., 2017), 而多数研究并没有发现情绪线索与IOR的交互作用(Berdica et al., 2017; Lange et al., 2008; Stoyanova et al., 2007)。这些研究认为情绪刺激不会对IOR产生影响, 支持IOR具有“盲目性”, 不会受到刺激性质干扰的观点。有研究者认为, 线索是以一种自动的外源性注意引起人们的关注, 这导致对线索类型的特征的忽视(Baijal & Srinivasan, 2011; Jia et al., 2019; Pérez-Dueñas et al., 2009, 2014; Silvert & Funes, 2016; 王敬欣 等, 2013), 所以当情绪刺激出现在线索化位置时不会受到IOR的影响。另一类研究将情绪刺激作为目标来探讨情绪刺激与IOR的交互作用。研究认为, 如果个体的注意被情绪刺激优先捕获从而使定位反应加快, 那么当它们出现在有效线索位置时会导致IOR效应会减少甚至消失(Silvert & Funes, 2016; 王敬欣 等, 2013)。Baijal和Srinivasan (2011)的研究中, 要求被试在检测到面部目标出现时进行按键反应, 结果发现负性情绪面孔目标削弱了IOR效应。Rutherford和Raymond (2010)的研究结果发现, 相比于中性刺激目标, 负性刺激目标产生了较少的IOR效应量。Silvert和Funes (2016)的研究采用面孔情绪效价辨别任务也发现了负性(恐惧)面孔比中性面孔的IOR效应量更少且出现的时间更晚。以上研究结果支持了IOR是一种“适应机制”, 会受到具有生物意义的情绪刺激的影响(Fox et al., 2002; Pérez-Dueñas et al., 2014; Taylor & Therrien, 2008; 王敬欣 等, 2013)。但也有一些研究结果发现, 采用辨别任务考察情绪刺激与IOR的加工过程时, 线索有效性和面孔效价在行为反应时和眼动结果上的交互作用均不显著(关荐 等, 2018), 且不同情绪效价的N170成分并没有受到IOR的影响(Jia et al., 2019)。对此, 研究者认为IOR发生在知觉层面, 是一种自下而上的加工, 但辨别视觉情绪效价的实验任务是一种自上而下的加工过程。由于二者处于不同的加工过程中, 因此不会产生相互的影响。
与单通道刺激相比, 视听双通道刺激整合后能够产生一种促进作用, 即冗余效应(redundant effect), 帮助被试准确且快速的对双通道刺激进行反应(Miller, 1986; Talsma & Woldorff, 2005)。此外, 视听双通道刺激发生视听觉整合后能够增强视听觉目标的感知觉显著性, 从而吸引个体的注意。这一特点有利于快速地检测、精准地识别以及有效地定位到相关信息(Stein & Stanford, 2008)。那么当发生整合后, 增强感知显著性的视听目标出现在曾经注意过的位置(产生IOR效应)时是否对IOR有影响, 这一问题引起了研究者的关注。van der Stoep等(2017)以视觉线索诱发不同目标通道条件下的IOR探讨了IOR是否会影响视听觉整合效应。以往研究表明相对多感觉反应增强(relative amount of multisensory response enhancement, rMRE)能够有效反映视听觉整合效应(Miller, 1986; Ulrich et al., 2007), 因此该研究通过比较有效线索和无效线索条件下的rMRE值来说明IOR对视听觉整合的影响。结果发现, 无效线索条件下的rMRE值显著高于有效线索条件, 这表明相比于无效线索条件, 有效线索条件下视觉线索引起的IOR降低了视听觉整合效应。此外, Tang等(2019)进一步操纵了基于通道的注意, 考察了单通道选择性注意和双通道分散性注意条件下视听觉整合效应对IOR的影响。结果发现, 在选择性注意条件下, 不同目标通道的IOR效应量无显著差异; 同时, rMRE值在线索有效性上也没有显著差异。但分散性注意条件下发现视听双通道目标存在IOR效应, 但效应量显著小于视觉通道目标产生的IOR效应量; 有效线索位置的rMRE值显著小于无效线索位置, 这表明在有效线索位置上比无效线索位置产生了更小的视听觉整合效应。之所以产生这样的结果是因为视听觉整合能够增强同时呈现的视觉刺激的知觉显著性(van der Burg et al., 2011), 而IOR是一种通过降低曾经注意过位置的知觉显著性来相应增强其他未注意位置显著性的机制(Klein, 2000)。因此, 当视听觉目标呈现在有效线索化位置时, 增强知觉显著性的视听觉目标可能会削弱IOR产生的抑制效应, 从而导致视听觉目标的IOR效应量小于视觉目标的IOR效应量(Tang et al., 2019)。综上, 以往的研究支持视听觉整合与IOR之间具有一定的交互作用。
当我们在日常交流中感知他人情绪时, 仅仅通过他人的面部表情或者声音表达很难准确地把握他人真实的情绪状态。但如果看到他人表情的同时听到他人的声音, 人们就能更加有效的体会他人的情绪, 从而促进人与人之间良好的交往。以往视听双通道呈现情绪刺激的研究结果也证实了, 整合加工后的视听双通道情绪一致刺激能够加快被试对情绪刺激的识别, 引起更大的情绪注意偏向(Klasen et al., 2014)。ERP研究结果发现, 当以视听双通道呈现情绪效价一致的刺激时, 会自动在早期100 ms内发生整合加工(Pourtois et al., 2000)。即使要求被试只注意一种通道的情绪, 视听双通道情绪一致刺激的整合也会发生(de Gelder & Vroomen, 2000)。也有研究发现, 当视听双通道呈现一致的视觉情绪和听觉情绪时, 比单通道诱发了波幅更小的N1和更大的P2波幅(Jessen & Kotz, 2011; Kokinous et al., 2015)。这些研究均表明, 视觉和听觉情绪一致刺激在早期知觉阶段就得到加工(李萍 等, 2019)。根据知觉抑制理论, IOR通过影响刺激的早期知觉加工来抑制对曾经搜索过的位置的注意, 这与视听双通道情绪一致刺激的整合加工发生的加工阶段相同。而以往采用经典的IOR范式的研究通常以单通道视觉形式呈现情绪刺激, 并且, 以往对情绪刺激与IOR关系的研究结果并不一致, 实验结果会受到情绪刺激呈现的位置、实验任务以及被试特性等因素的影响。因此, 本研究采用线索-目标范式, 并将视听双通道情绪一致刺激作为目标进一步地考察情绪刺激与IOR在同一加工阶段时, 情绪刺激对IOR的影响。研究分为两个实验, 采用线索-目标范式考察视听双通道情绪一致刺激对IOR的影响。实验1探讨视听双通道情绪一致刺激对IOR的影响。实验1假设, 由于视听双通道情绪一致刺激与IOR的加工阶段相同, 此时二者之间会产生注意资源的竞争, 而情绪刺激会占用更多的加工资源, 所以视听双通道情绪一致刺激能够影响IOR, 即视听双通道条件下情绪一致刺激能够削弱IOR效应。此外, 以往视听目标IOR效应的研究发现当同时呈现非情绪性的听觉刺激时, 也会发生视听觉整合, 促进视觉加工, 从而影响IOR的产生(Tang et al., 2019; van der Stoep et al., 2017)。那么视听双通道情绪一致刺激对IOR的影响是否是由听觉通道一致的情绪刺激导致的, 即是否对听觉通道情绪刺激进行了加工。因此, 为了进一步考察视听双通道情绪一致刺激对IOR的影响, 本研究在实验1的基础上进行了实验2, 仅操纵听觉通道的情绪刺激效价与视觉通道的情绪刺激效价不一致, 其他条件与实验1相同。实验2假设视听双通道呈现不一致的情绪刺激时, 视听双通道情绪不一致刺激与IOR之间不存在交互作用。
2 实验1:视听双通道情绪一致对返回抑制的影响
2.1 方法
2.1.1 被试
本实验在苏州大学随机招募33名被试, 被试的听力, 视力或矫正视力正常, 习惯手均为右手, 未有精神疾病, 并报告未参加过同类实验, 于实验结束后获取相应的报酬。其中有2名被试正确率低于80%, 低正确率的出现可能是由被试不认真或注意力不集中导致, 因此后续的数据分析中排除2名低正确率的被试, 共31名被试(男生15名, 女生16名, 平均年龄19.97 ± 2.06岁)纳入后续的数据分析。为了评估本研究的统计检验力, 通过软件G*Power 3.1 (Faul et al., 2009; Faul et al., 2007)对双侧配对样本t检验进行了敏感性分析(sensitivity analysis), 设置α = 0.05, power = 0.80, 计算出effect size dz = 0.52, 根据以往研究本研究的效应量达到了中等效应量, 因此本研究的统计检验力较好。
2.1.2 实验仪器与材料
整个实验过程在隔音且较暗的室内环境里实施。所有的刺激呈现在23英寸的Dell-3020MT型号显示器上, 分辨率为1024 × 768, 刷新率为60 Hz。实验使用白色背景, 其中每个矩形框的水平视角为2.34°, 垂直视角为3.36°, 三个矩形框的视角范围约为 ± 5°, 实验过程中的中央注视点的水平和垂直视角均为0.5°, 线索是空心圆环, 中央线索是黑色圆环, 水平和垂直视角均为2°。实验过程中保持被试的双眼距离显示器屏幕中央约为65 cm。实验在E-prime 2.0中完成程序的编写、呈现和数据归纳记录。
有研究认为, 哺乳动物存在一个进化的恐惧系统, 以快速检测存在社会威胁的面部表情, 即与其它信号相比, 人类大脑更容易识别到恐惧刺激(Anderson et al., 2003; Mineka & Öhman, 2002; Troiani et al., 2014; Vizueta et al., 2012)。且以往采用线索-目标范式来探讨情绪与返回抑制关系的研究也多采用恐惧情绪面孔与中性情绪进行对比, 如 Silvert和Funes (2016)采用线索-目标范式, 并要求被试辨别靶子位置呈现的是恐惧面孔还是中性面孔来探讨视觉情绪刺激与返回抑制的关系, 结果发现负性(恐惧)面孔比中性面孔的IOR效应量更少且出现的时间更晚。因此, 本研究选择恐惧面孔与中性面孔作为情绪刺激材料。
视觉材料:选自NimStim刺激库(Tottenham et al., 2009)中4个人的面孔表情图片, 每个人包括负性(恐惧面孔, 效价:3.37 ± 0.33分; 唤醒度:5.70 ± 0.32分)、中性(平静面孔, 效价:4.57 ± 0.19分; 唤醒度3.45 ± 0.29分)两种面孔表情图片, 共有8张图片, 男女比例为50%, 并将图片亮度、对比度和尺寸在Adobe PhotoShop 2019图像处理软件中调整统一。
听觉材料:选自蒙特利尔情绪声音刺激库(Belin et al., 2008)中4个人的情绪声音, 每个人包括负性(恐惧声音, 效价:3.36 ± 0.28分; 唤醒度:5.30 ± 0.68分)、中性(平静声音, 效价:4.54 ± 0.25分, 唤醒度:3.50 ± 0.24分)两种情绪声音, 共有8个声音, 男女比例为50%, 之后经Adobe Audition CC软件进行处理, 截取声音时长为1000 ms, 采样率为44100 Hz, 并将每个声音都剪辑成左声道和右声道两种, 被试只能从单耳听到每个声音。声音刺激通过头戴式耳麦(ATH-WS99)呈现, 音量均控制在65 dB。
2.1.3 实验设计与流程
实验采用2 (线索有效性:有效vs.无效) × 2 (刺激通道:视觉vs.视听觉) × 2 (情绪效价:负性vs.中性)的被试内设计, 因变量为反应时和正确率。实验中视听双通道情绪呈现是指在显示器屏幕上看到视觉情绪面孔的同时, 同侧耳麦会播放相应的情绪效价一致的声音情绪(视觉负性面孔-听觉负性声音; 视觉中性面孔-听觉中性声音)。
采用线索-目标范式, 并在线索和目标中间加入了中央重定向事件(中央线索)诱发更稳定的IOR现象, SOA在1000~1100 ms内变化(Prime & Ward, 2006; Taylor & Therrien, 2008)。每个试次的实验流程如图1所示:开始前屏幕中出现3个矩形框, 中央注视点“+”呈现在中间矩形框中。750~850 ms后一个空心的圆环以相等的概率出现在两边矩形框中的一个作为外周线索, 呈现时长200 ms。在300 ms间隔后, 中央注视点“+”被黑色圆环替换(中央线索), 经过200 ms后又变回中央注视点“+”。之后间隔300~400 ms, 呈现目标刺激(视觉情绪面孔vs.视觉情绪面孔-听觉情绪声音), 目标刺激以相等的概率出现在左边或右边的矩形框中, 要求被试在目标呈现后辨别视觉面孔情绪效价, 同时尽可能快速准确地做出按键反应, 中性按键盘F键, 负性按键盘J键(反应键在另外一半被试中进行了平衡, 即中性按键盘J键, 负性按键盘F键), 按键后目标刺激消失(刺激最长呈现时间为1000 ms), 并且要求被试在实验全程盯住中央注视点“+”。
图1
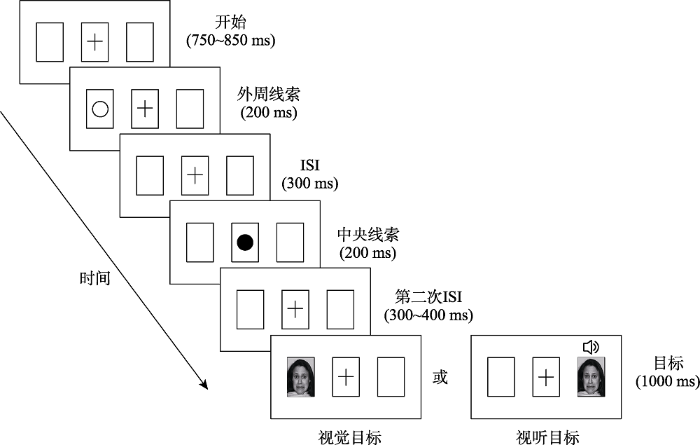
实验由练习实验和正式实验两个部分组成, 练习实验有64个试次, 用于被试循环练习来熟悉明白实验程序。正式实验中有576个试次, 其中包含64个捕获试次(无任何目标刺激出现), 分为4个block进行, 每个block有144个试次。实验中, 中性和负性刺激(1 : 1), 视觉单通道和视听双通道(1 : 1)以及有效线索、无效线索(1 : 1)均随机出现。被试可以在两个block间短暂休息1分钟, 整个实验时长约为60分钟。
2.2 结果与分析
2.2.1 正确率
对正确率进行2 (线索有效性:有效线索vs.无效线索) × 2 (通道:视觉vs.视听觉) × 2 (情绪效价:负性vs.中性)的重复测量方差分析, 结果发现, 线索有效性主效应不显著, F(1, 30) = 2.04, p = 0.16; 通道主效应显著, F(1, 30) = 7.81, p = 0.01, ηp2 = 0.21, 即单通道视觉目标的正确率(97%)显著低于视听双通道目标(98%), 体现了双通道加工的优势; 情绪效价主效应显著, F(1, 30) = 7.55, p = 0.01, ηp2 = 0.20, 即辨别负性目标的正确率(97%)要显著小于中性目标(98%); 线索有效性与通道的交互作用不显著, F < 1; 线索有效性与情绪效价的交互作用不显著, F < 1; 通道与情绪效价的交互作用显著, F(1, 30) = 47.58, p < 0.001, ηp2 = 0.61; 线索有效性、通道和情绪效价三因素交互作用不显著, F(1, 30) = 3.51, p = 0.07。
2.2.2 反应时
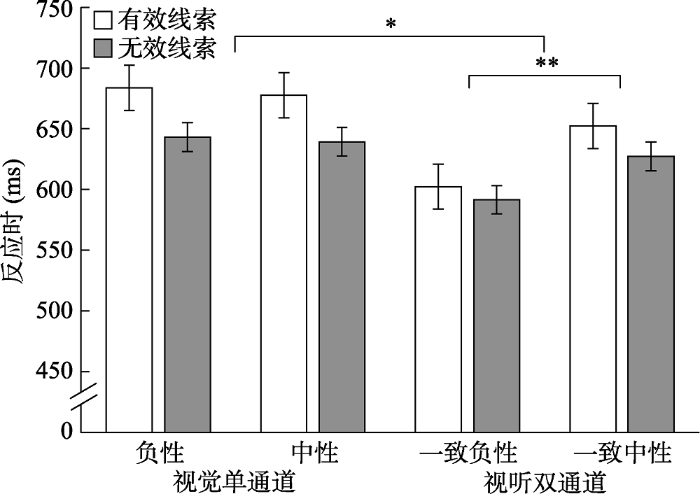
剔除判断错误的试次; 剔除小于200 ms和大于2000 ms的试次; 剔除平均数正负3个标准差之外的试次。对反应时进行2 (线索有效性:有效线索vs.无效线索) × 2 (通道:视觉vs.视听觉) × 2 (情绪效价:负性vs.中性)的重复测量方差分析(图2)。结果表明线索有效性的主效应显著, F(1, 30) = 92.04, p < 0.001, ηp2 = 0.75, 有效条件下的反应(654 ms)显著慢于无效条件下的反应(625 ms), 即出现稳定的IOR现象; 通道的主效应显著, F(1, 30) = 38.93, p < 0.001, ηp2 = 0.57, 即单通道视觉条件下的反应(661 ms)显著慢于视听双通道条件下反应(618 ms), 产生冗余效应, 体现了双通道加工的优势; 情绪效价主效应显著, F(1, 30) = 7.72, p = 0.01, ηp2 = 0.21, 即辨别负性目标的反应(630 ms)显著快于中性目标的反应(649 ms), 表明情绪刺激引起了注意偏向。线索有效性和情绪效价的交互作用显著, F(1, 30) = 4.28, p = 0.047, ηp2 = 0.13; 通道和情绪效价的交互作用显著, F(1, 30) = 55.62, p < 0.001, ηp2 = 0.65; 线索有效性和通道的交互作用显著, F(1, 30) = 32.17, p < 0.001, ηp2 = 0.52。进一步简单效应分析表明, 视觉单通道条件下, 无效线索条件下的反应(641 ms)显著快于有效线索条件(681 ms), F(1, 30) = 103.49, p < 0.001, ηp2 = 0.78; 视听双通道条件下, 无效线索条件下的反应(609 ms)也显著快于有效线索条件(627 ms), F(1, 30) = 31.57, p < 0.001, ηp2 = 0.51, 即在两种通道条件下IOR现象均稳定出现。线索有效性、通道和情绪效价三者的交互作用显著, F(1, 30) = 6.62, p = 0.02, ηp2 = 0.18。
图2
2.2.3 IOR效应量
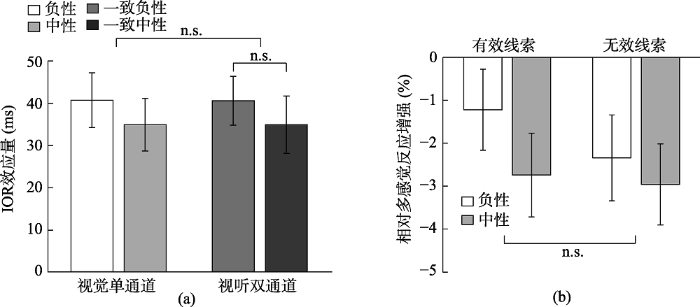
为了考察不同通道类型和情绪效价如何对IOR效应产生影响, 以IOR效应量(线索有效条件下的反应时减去线索无效条件下的反应时)为指标, 对数据进行了2 (通道:视觉vs.视听觉) × 2 (情绪效价:负性vs.中性)的重复测量方差分析(图3a)。结果发现通道主效应显著, F(1, 30) = 32.16, p < 0.001, ηp2 = 0.52, 视听双通道条件下的IOR效应量(18 ms)显著小于单通道视觉的IOR效应量(40 ms); 情绪效价的主效应也显著, F(1, 30) = 4.27, p = 0.047, ηp2 = 0.13, 负性目标下的IOR效应量(26 ms)显著小于中性目标的IOR效应量(32 ms)。通道与情绪目标的交互作用显著, F(1, 30) = 6.62, p = 0.02, ηp2 = 0.18, 简单效应分析表明, 在视听双通道条件下, 辨别负性目标的IOR效应量(11 ms)显著小于中性目标(25 ms), F(1, 30) = 8.12, p = 0.008, ηp2 = 0.21。但单通道条件下, 辨别负性目标(41 ms)和中性目标(38 ms)之间的IOR效应量无显著差异, F < 1。此外, 在辨别负性目标条件下, 视听双通道的IOR效应量(11 ms)显著小于视觉单通道的IOR效应量(41 ms), F(1, 30) = 38.82, p < 0.001, ηp2 = 0.56; 辨别中性目标条件下, 视听双通道的IOR效应量(25 ms)显著小于视觉单通道的IOR效应量(38 ms), F(1, 30) = 6.75, p = 0.01, ηp2 = 0.18。以上结果说明仅在视听双通道条件下情绪刺激显著地影响IOR效应, 并且无论情绪效价如何, 视听双通道比视觉单通道更能削弱IOR效应量。
图3
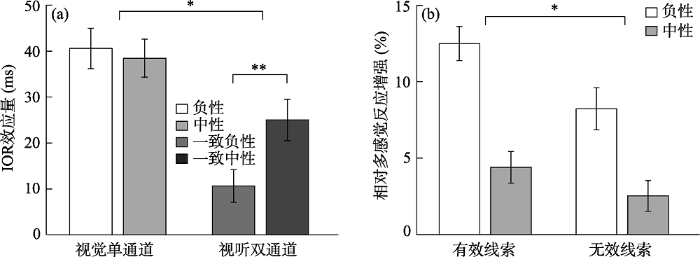
图3
(a)各个条件下的IOR效应量(RT有效–RT无效), (b) 各个条件下的相对多感觉反应增强(rMRE) (注:*p < 0.05, **p < 0.01, ***p < 0.001)
2.2.4 相对多感觉反应增强(rMRE)
$\text{rMRE=}\frac{\text{median}(\text{R}{{\text{T}}_{\text{V}}})-\text{median}(\text{R}{{\text{T}}_{\text{AV}}})}{\text{median}(\text{R}{{\text{T}}_{\text{V}}})}\times \text{100 }\!\!%\!\!\text{ }$
首先对各个条件下的rMRE进行单样本t检验(与0进行比较), 若rMRE值显著大于0, 说明多感觉刺激加速量显著大于零, 即出现了多感觉反应增强效应, 反之则没有出现。结果发现有效线索负性情绪(t (30) = 11.16, p < 0.001, Cohen's d = 2.00, 95% CI = [10.20, 14.78])、有效线索中性情绪(t (30) = 5.99, p < 0.001, Cohen's d = 1.08, 95% CI = [5.42, 11.03])、无效线索负性情绪(t (30) = 4.243, p < 0.001, Cohen's d = 0.76, 95% CI = [2.28, 6.51])、无效线索中性情绪(t (30) = 2.51, p = 0.018, Cohen's d = 0.45, 95% CI = [0.47, 4.58])的结果均显著大于0, 对于视听双通道目标的反应时要比单通道视觉目标的反应时更短, 产生冗余效应, 表明存在双通道加工优势。然后对rMRE进行2 (线索有效性:有效线索vs.无效线索) × 2 (情绪效价:负性vs.中性)的重复测量方差分析(图3b)。结果发现线索有效性的主效应显著, F(1, 30) = 22.61, p < 0.001, ηp2 = 0.43, 有效线索位置上的rMRE (8.44%)显著大于无效线索位置(5.38%)。情绪效价的主效应显著, F(1, 30) = 59.47, p < 0.001, ηp2 = 0.67, 负性目标的rMRE (10.36%)显著大于中性目标(3.46%)。线索有效性与情绪效价的交互作用显著, F(1, 30) = 5.15, p = 0.03, ηp2 = 0.15, 进一步简单效应分析表明, 在有效线索条件下, 负性目标的rMRE (12.49%)显著大于中性目标(4.40%), F(1, 30) = 75.56, p < 0.001, ηp2 = 0.72; 在无效线索条件下, 负性目标的rMRE (8.23%)显著大于中性目标(2.53%), F(1, 30) = 25.13, p < 0.001, ηp2 = 0.46。在负性目标条件下, 有效线索位置的rMRE (12.49%)显著大于无效线索位置的rMRE (8.23%), F(1, 30) = 26.77, p < 0.001, ηp2 = 0.47, 在中性目标条件下, 有效线索位置的rMRE (4.40%)显著大于无效线索位置的rMRE (2.53%), F(1, 30) = 4.90, p = 0.04, ηp2 = 0.14。
2.2.5 讨论
实验1的结果发现, 被试在有效线索化条件下的反应均慢于无效线索化条件反应, 表明IOR现象稳定出现。单通道视觉条件下情绪刺激与IOR交互作用不显著(图3a), 但视听双通道条件下情绪一致刺激与IOR的交互作用显著, 且一致负性目标的IOR效应量显著小于一致中性目标的IOR效应量, 这表明发生在早期知觉阶段的视听双通道情绪一致刺激能够调节IOR效应, 与实验假设相符。为了验证本研究中视听整合是否发生, 计算了rMRE值(图3b)。结果发现, 视听双通道条件下的确发生了整合现象, 而且一致负性目标的整合效应显著大于一致中性目标。因此实验1表明, 当在经典的IOR范式的目标位置上以视听双通道呈现一致的情绪刺激时, 视听双通道情绪一致刺激与IOR均处于早期的知觉阶段, 由于二者之间会产生注意资源的竞争, 情绪刺激会占用更多的加工资源, 因此具有注意优先性的视听双通道情绪一致刺激能够削弱IOR效应。但以往视听目标IOR效应的研究发现, 当同时呈现非情绪性的听觉刺激时, 也会发生视听觉整合, 促进视觉加工, 从而影响IOR效应(Tang et al., 2019; van der Stoep et al., 2017)。那么实验1中视听双通道情绪一致刺激对IOR的影响是否是由听觉通道一致的情绪刺激导致的, 即是否对听觉通道情绪刺激进行了加工还有待确定。因此本研究设计了实验2, 操纵听觉通道的情绪刺激效价与视觉通道的情绪刺激效价不一致, 其他条件与实验1相同, 进一步探讨视听双通道情绪一致刺激对IOR的影响。
3 实验2:视听双通道情绪不一致对返回抑制的影响
3.1 方法
3.1.1 被试
本实验在苏州大学随机招募32名被试, 被试的听力, 视力或矫正视力正常, 习惯手均为右手, 未有精神疾病, 并报告未参加过同类实验, 于实验结束后获取相应的报酬。其中有1名被试正确率低于80%, 低正确率的出现可能是由被试不认真或注意力不集中导致, 因此后续的数据分析中排除1名低正确率的被试, 共31名被试(男生13名, 女生18名, 平均年龄20.75 ± 2.31岁)纳入后续的数据分析。为了评估本研究的统计检验力, 通过软件G*Power 3.1 (Faul et al., 2009; Faul et al., 2007)对双侧配对样本t检验进行了敏感性分析(sensitivity analysis), 设置α = 0.05, power = 0.80, 计算出effect size dz = 0.52, 根据以往研究本研究的效应量达到了中等效应量, 因此本研究的统计检验力较好。
3.1.2 实验仪器与材料
同实验1。
3.1.3 实验设计与流程
与实验1的不同在于目标以视听双通道呈现时, 视听双通道呈现的情绪效价是不一致的(视觉负性面孔-听觉中性声音; 视觉中性面孔-听觉负性声音), 除此以外与实验1相同。
3.2 结果与分析
3.2.1 正确率
对正确率进行2 (线索有效性:有效线索vs.无效线索) × 2 (通道:视觉vs.视听觉) × 2 (情绪效价:负性vs.中性)的重复测量方差分析。结果发现, 线索有效性的主效应不显著, F(1, 30) = 1.38, p = 0.25; 通道主效应不显著, F(1, 30) = 3.90, p = 0.06; 情绪效价主效应显著, F(1, 30) = 15.07, p = 0.001, ηp2 = 0.33, 即辨别负性目标的正确率(97%)要显著小于中性目标(98%); 线索有效性与通道的交互作用不显著, F < 1; 线索有效性与情绪效价的交互作用不显著, F < 1; 通道与情绪效价的交互作用显著, F(1, 30) = 6.73, p = 0.02, ηp2 = 0.18; 线索有效性、通道和情绪效价三因素交互作用不显著, F < 1。
3.2.2 反应时
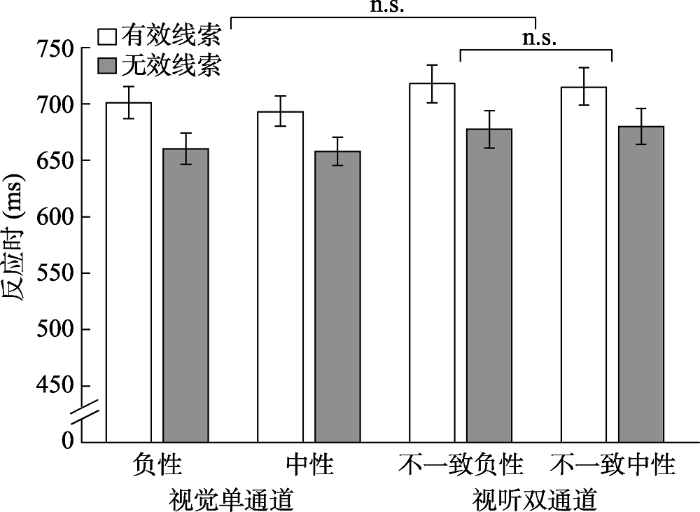
剔除判断错误的试次; 剔除小于200 ms和大于2000 ms的试次; 剔除平均数正负3个标准差之外的试次。对反应时进行2 (线索有效性:有效线索vs.无效线索) × 2 (通道:视觉vs.视听觉) × 2 (情绪效价:负性vs.中性)的重复测量方差分析(图4)。结果发现线索有效性的主效应显著, F(1, 30) = 60.07, p < 0.001, ηp2 = 0.67, 线索有效条件下的反应(707 ms)显著慢于无效条件下的反应(669 ms), 即出现稳定的IOR现象; 通道的主效应显著, F(1, 30) = 9.07, p = 0.01, ηp2 = 0.23, 即单通道视觉条件下的反应(678 ms)显著快于视听双通道条件下反应(698 ms), 以往视听双通道加工优势没有出现, 当视听双通道呈现刺激不一致时比单通道具有更长的反应时; 情绪效价主效应不显著, F < 1。线索有效性和通道的交互作用不显著, F < 1; 线索有效性和情绪效价的交互作用不显著, F(1, 30) = 2.17, p = 0.15; 通道和情绪效价的交互作用不显著, F < 1; 线索有效性、通道和情绪效价三者的交互作用不显著, F < 1。
图4
3.2.3 IOR效应量
以IOR效应量为指标, 对数据进行2 (通道:视觉vs.视听觉) × 2 (情绪效价:负性vs.中性)的重复测量方差分析。结果如图5(a)所示, 通道主效应不显著, F < 1; 情绪效价主效应不显著, F(1, 30) = 2.17, p = 0.15。通道与情绪效价的交互作用不显著, F < 1。
图5
3.2.4 相对多感觉反应增强(rMRE)
首先对各个条件下的rMRE进行单样本t检验(与0进行比较), 发现有效线索负性情绪条件下没有显著小于0 (t(30) = -1.31, p = 0.20), 在无效线索负性情绪(t (30) = -2.86, p = 0.008, Cohen's d = -0.51, 95% CI = [-4.69, -0.78])、有效线索中性情绪(t (30) = -2.38, p = 0.024, Cohen's d = -0.43, 95% CI = [-4.35, -0.33])和无效线索中性情绪(t (30) = -3.19, p = 0.003, Cohen's d = -0.57, 95% CI =[-4.85, -1.06])条件下结果均显著的小于0, 表明视听双通道呈现不一致刺激时没有表现出冗余效应, 双通道加工优势不存在。然后对rMRE进行2 (线索有效性:有效线索vs.无效线索) × 2 (情绪效价:负性vs.中性)的重复测量方差分析。结果如图5(b)所示, 线索有效性的主效应不显著, F(1, 30) = 1.95, p = 0.17; 情绪效价主效应不显著, F(1, 30) = 1.09, p = 0.31。线索有效性与情绪效价的交互作用不显著, F < 1。
3.2.5 讨论
实验2的结果显示, 在不同条件下IOR效应均稳定出现, 但实验1在视听双通道条件下发现的情绪刺激与IOR的交互作用在实验2中没有发现。同时, 不同通道条件下的IOR效应量无显著差异, 即不一致的视听双通道情绪刺激没有对IOR产生影响。从rMRE值(图5b)的结果发现, 当视听双通道呈现不一致的情绪刺激没有出现冗余效应, 被试对双通道不一致条件下的反应不具有加工优势。根据实验2的结果可以推断, 不一致的视听双通道情绪刺激可能未在知觉阶段发生整合加工, 此时与IOR处于不同的加工阶段(知觉阶段), 因此不会削弱IOR效应。由此推断, 实验1中视听双通道情绪一致刺激对IOR的影响是由听觉通道一致的情绪刺激导致的, 即对听觉通道的情绪刺激进行了加工。综上, 实验2的结果表明, 视听双通道情绪不一致刺激未在知觉阶段发生整合加工时, 不会影响知觉阶段发生的IOR效应, 这与单通道条件下情绪刺激与IOR的结果一致(Jia et al., 2019)。
4 讨论
本研究采用线索-目标范式, 操纵线索有效性、目标情绪效价和目标通道三个变量, 考察视听双通道情绪一致刺激对IOR的影响。实验1视听双通道呈现一致的情绪刺激时发现无效线索条件下的反应快于有效线索, 即IOR现象稳定的出现; 同时出现视听双通道的冗余效应, 视听双通道反应快于视觉单通道; 此外, 与以往研究一致的情绪注意偏向出现, 即被试对负性目标的反应显著快于中性目标。进一步分析发现, 视听双通道条件下情绪一致刺激与IOR的交互作用, 具体表现为, 在视听双通道条件下, 一致的负性目标产生的IOR效应量要显著地小于中性目标的IOR效应量; 但在视觉单通道条件下并没有发现二者的交互作用。此外, 视听双通道目标产生的IOR效应量要显著地小于视觉单通道的IOR效应量。实验1的结果表明, 当视听双通道呈现一致的情绪刺激时能够削弱发生在同一加工阶段的IOR效应。在实验2中, 当以视听双通道呈现不一致的情绪刺激时, 结果发现视觉单通道与视听双通道均产生了稳定的IOR效应, 但视听双通道条件下情绪不一致刺激与IOR之间不存在交互作用, 同时视觉单通道IOR效应量与视听双通道IOR效应量也不存在显著差异。实验2的结果表明, 呈现视听双通道情绪不一致刺激时对IOR没有影响。综合实验1与实验2的结果表明, 视听双通道呈现情绪一致刺激时, 才会影响同一阶段的IOR效应。
实验1发现视觉单通道条件下情绪效价(负性vs.中性)产生的IOR效应量不存在显著差异。这与以往研究结果一致, 表明视觉单通道呈现情绪刺激时并不会影响辨别任务中的IOR效应, 进一步支持了IOR的确具有“盲目性”, 不会受到情绪刺激干扰影响的观点(Berdica et al., 2017; Jia et al., 2019)。研究者认为IOR是在知觉层面发生的注意抑制现象, 是一种自下而上的行为过程。而在本研究中, 实验任务是要求被试辨别目标刺激的情绪效价, 这是一种经过深度加工以后才可以完成的任务, 属于对情绪刺激的意识反应阶段, 在辨别刺激属性时会受到多种因素的影响, 例如相关的个人经验等, 这是一种自上而下的加工过程(Jia et al., 2019)。由于两个不同的加工过程沿着不同的神经通路独立发生, 因此二者之间不会产生相互影响。这一研究结果同时也进一步支持了IOR的确发生在知觉阶段。
重要的是, 实验1采用了在知觉阶段发生整合的视听双通道情绪一致刺激进一步探究情绪刺激与IOR的关系。结果发现了视听双通道条件下情绪一致刺激与IOR之间的交互作用, 即视听双通道呈现一致的负性目标比一致的中性目标产生了更小的IOR效应量(图3a)。结合反映整合效应的rMRE值发现, 在视听双通道条件下, 一致负性目标和一致中性目标都产生了视听觉整合效应(图3b), 被试对视听双通道目标的反应显著快于视觉单通道目标, 视听双通道的加工优势出现, 即产生冗余效应。此外, 本研究发现, 无论是在有效线索位置还是无效线索位置, 一致负性目标的rMRE值都比中性目标大, 即一致负性目标在知觉阶段发生了更显著的整合效应(图3b)。这一结果与以往视听双通道情绪的研究一致, 相比于中性目标, 当视听双通道呈现一致的负性目标(如恐惧)时, 更能够促进整合过程, 且早期整合效应更强, 这同样反应了情绪信息因具有进化意义而表现出双通道情绪整合的情绪特异性(Lang et al., 2000)。因此, 研究结果证明了发生在早期知觉阶段的视听双通道情绪一致刺激与IOR处于同一加工阶段, 情绪刺激能够削弱IOR效应, 说明IOR具有一定的适应性, 能够帮助人们更好地适应社会, 与实验假设相符。根据IOR的知觉抑制理论, IOR发生在早期知觉阶段, 通过对先前搜索的位置进行注意抑制来促进搜索效率(Prime & Jolicœur, 2009; Reuter-Lorenz et al., 1996)。但由于注意资源的有限性, IOR会受到其他能够引起注意偏向的刺激的影响, 从而适应环境变化。以往相关研究的结果表明, 当情绪刺激以视听双通道呈现时, 视听双通道情绪一致刺激的整合加工也发生在知觉阶段(de Gelder & Vroomen, 2000; Jessen & Kotz, 2011; Kokinous et al., 2015; Pourtois et al., 2000)。综上, 本研究结果证明了情绪刺激的加工与IOR的产生同在知觉阶段时, 由于二者处于竞争状态, 情绪刺激会占用更多的加工资源, 所以具有注意优先性的情绪刺激能够削弱IOR效应(Pan et al., 2017), 这也进一步支持了IOR的知觉抑制理论。
此外, 实验1发现视听双通道目标比视觉单通道目标更能削弱IOR效应量, 无论是在负性目标还是中性目标条件下, 不同通道的IOR效应量都有显著差异(图3a)。以往研究表明, 当以视听双通道呈现目标刺激时, 视觉和听觉通道会因为时间和空间的一致性发生整合。为了反映整合效应, 实验1在数据分析时计算了rMRE值。由图3b可知, 实验1中的确发生了视听整合(各种条件下rMRE值均显著大于0), 并且有效线索位置的整合效应显著的大于无效线索位置。以往视听目标IOR效应的研究发现同时呈现视听刺激时能够发生视听觉整合现象, 从而对IOR有一定的影响, 研究表明当视听觉目标 IOR (降低显著性)遇到视听觉整合(增加感知觉显著性)时, 视听觉整合可以帮助视听觉目标抵抗其在早期感知/注意处理中被抑制(Tang et al., 2019)。也就说当刺激以视听这样的双通道形式出现时, 会发生视听觉整合现象, 而这样的整合由于比单通道能更加增强刺激感知显著性从而促进对刺激的加工(Stein & Stanford, 2008)。因此当视听双通道目标出现在了曾经注意过的位置时, 相比于单通道它会因为增强的感知显著性去削弱IOR的抑制效应。这与本研究的结果一致, 即视听双通道目标比视觉单通道目标更能削弱IOR效应。
实验1发现视听双通道情绪一致条件下削弱IOR, 同时也发现了不同通道间的IOR效应量差异。在目标位置呈现视听刺激的IOR研究发现当同时呈现非情绪性的听觉刺激时, 也会发生视听觉整合, 促进视觉加工, 从而影响IOR效应(Tang et al., 2019; van der Stoep et al., 2017)。因此, 实验2通过设置视听双通道呈现不一致的情绪刺激进一步探究视听双通道情绪一致刺激对IOR的影响。结果发现, 实验2结果与实验1不同。在实验2中视听双通道情绪不一致刺激与IOR不存在交互作用; 同时, 不同通道条件下的IOR效应量无显著差异。实验2与实验1唯一不同在于听觉通道的情绪刺激效价与视觉通道情绪刺激效价是否一致, 其他条件保持不变。因此可以推断, 视听双通道情绪一致刺激对IOR的影响是由听觉通道一致的情绪刺激导致的, 即对听觉通道的情绪刺激进行了加工, 而视听双通道情绪不一致刺激可能未在知觉阶段发生整合, 此时与IOR处于不同的加工阶段, 因此不会削弱IOR效应。此外, 实验2中rMRE值(图5b)的结果发现, 当视听双通道呈现不一致的情绪刺激时, 视听双通道条件下没有出现冗余效应, 被试对视听双通道不一致条件下的反应不具有加工优势, 甚至实验2中要求对视觉通道情绪效价进行辨别时, 受到了不一致的听觉通道情绪声音的干扰, 具体表现为视听双通道条件下的反应比视觉单通道条件下更慢(图4)。这一结果与以往视听双通道情绪的研究结果一致, 即对一种通道的情绪辨别会被其他通道呈现一致的情绪刺激所增强, 但会被其他通道呈现不一致的情绪刺激所削弱(Collignon et al., 2008; de Gelder & Vroomen, 2000; Müller et al., 2011)。以往对Stroop效应(视觉颜色)与IOR间关系的研究发现两者在视觉通道下交互作用显著, 并且线索化位置的干扰量减小, 这可能是由于IOR抑制了前反应水平的信息加工(Chen et al., 2006)。Zhao等人(2017)采用词-面孔Stroop范式探讨了视觉通道下情绪干扰效应与IOR之间的关系, 结果发现情绪干扰与IOR也发生了交互作用。然而, 本研究结果发现, 双通道不一致情绪刺激和单通道视觉情绪刺激没有与线索有效性产生交互作用, 即听觉不一致情绪的干扰效应没有影响IOR效应。这可能由于视听双通道呈现不一致的情绪刺激时未在早期知觉阶段发生整合, 而返回抑制是通过影响刺激的早期知觉加工产生抑制效应, 所以二者没有产生交互作用, 这进一步地支持了实验1的结果, 表明视听双通道情绪一致刺激削弱IOR是因为呈现一致的视听双通道情绪刺激时会影响同一阶段(知觉阶段)的IOR效应, 从而进一步支持了IOR的确发生在知觉阶段。
5 结论
(1)当以视听双通道呈现一致的情绪刺激时, 由于一致的情绪刺激在知觉阶段发生整合, 从而影响了发生在同一知觉阶段的IOR效应;
(2)当以视听双通道呈现不一致的情绪刺激时, 视听双通道情绪不一致刺激对IOR没有影响。
当视听双通道情绪刺激一致时, 才会在知觉阶段发生整合进而削弱IOR效应量, 研究结果进一步支持了IOR的知觉抑制理论。
参考文献
Neural correlates of the automatic processing of threat facial signals
DOI:10.1523/JNEUROSCI.23-13-05627.2003 URL [本文引用: 1]
Lesions of the human amygdala impair enhanced perception of emotionally salient events
DOI:10.1038/35077083 URL [本文引用: 1]
Emotional and hemispheric asymmetries in shifts of attention: An ERP study
DOI:10.1080/02699931.2010.492719 URL [本文引用: 2]
The Montreal Affective Voices: A validated set of nonverbal affect bursts for research on auditory affective processing
DOI:10.3758/BRM.40.2.531 URL [本文引用: 1]
A comprehensive look at phobic fear in inhibition of return: Phobia-related spiders as cues and targets
DOI:S0005-7916(16)30032-5
PMID:27517673
[本文引用: 3]

The so called inhibition of return (IOR) effect refers to a bias against returning attention to a location which was previously investigated. Because emotionally salient material has the capacity to capture and hold attention it has been suggested that this material may disrupt this otherwise impressively stable phenomenon.40 students participated in the experiment. Black and white schematic drawings of a spider, a butterfly or a cross were used as cues. A black dot, a spider, a butterfly or a cross were used as targets. Participants were required to press a key whenever the target picture appeared. Subsequently, they rated the pictures on valence and arousal.Results showed that the IOR effect remained stable and did not diminish with either fear-related cues or fear-related targets. This data adds strong arguments for the stability of IOR.The spider fearful participants were not diagnosed patients. They still meet the criteria for spider fear but follow-up studies should pursue the same question with a specific focus on participants' levels of anxiety.This study is a contribution to the debate on how emotions affect or do not affect attentional processes such as the IOR. IOR appears to be a robust phenomenon and the emotional valence of neither the cue nor the emotional valence of the target can override it.Copyright © 2016 Elsevier Ltd. All rights reserved.
Distinct neural correlates for resolving stroop conflict at inhibited and noninhibited locations in inhibition of return
It is well documented that the anterior cingulate cortex (ACC) and the dorsolateral prefrontal cortex (DLPFC) are intensively involved in conflict control. However, it remains unclear how these "executive" brain regions will act when the conflict control process interacts with spatial attentional orienting. In the classical spatial cueing paradigm [Posner, M. I., & Cohen, Y. (1984). Components of visual orienting. In H. Bouma & D. G. Bouwhuis (Eds.), Attention and performance X (pp. 531-556). Hillsdale, NJ: Erlbaum], response to a target is delayed when it appears at the cued location compared with at the uncued location, if the time interval between the cue and the target is greater than 300 msec. This effect of inhibition of return (IOR) can alter the resolution of Stroop conflict such that the Stroop interference effect disappears at the cued (inhibited) location [Vivas, A. B., & Fuentes, L. J. Stroop interference is affected in inhibition of return. Psychonomic Bulletin and Review, 8, 315-323, 2001]. In this event-related functional magnetic resonance study, we investigate the differential neural mechanisms underlying interactions between pre-response interference, response interference, and spatial orienting. Two types of Stroop words [incongruent response-eligible words (IE), incongruent response-ineligible words (II)] and neutral words were presented either at the cued or uncued location. The significant pre-response interference at the uncued location activated the left rostral ACC as compared with at the cued location. Moreover, although the IE words which have conflicts at both pre-response and response levels did not cause significant behavioral interference at the cued location, they activated the left DLPFC as compared with at the uncued location. Furthermore, neutral words showed significant IOR effects behaviorally, and they activated the left frontal eye field (FEF) at the uncued location relative to the cued location. These results suggest that the left rostral ACC is involved in the interaction between pre-response conflict and IOR, whereas the left DLPFC is involved in the interaction between response conflict and IOR. Moreover, the FEF is involved in shifting attentional focus to novel locations during spatial search.
Audio-visual integration of emotion expression
DOI:10.1016/j.brainres.2008.04.023
PMID:18495094
[本文引用: 1]
Regardless of the fact that emotions are usually recognized by combining facial and vocal expressions, the multisensory nature of affect perception has scarcely been investigated. In the present study, we show results of three experiments on multisensory perception of emotions using newly validated sets of dynamic visual and non-linguistic vocal clips of affect expressions. In Experiment 1, participants were required to categorize fear and disgust expressions displayed auditorily, visually, or using congruent or incongruent audio-visual stimuli. Results showed faster and more accurate categorisation in the bimodal congruent situation than in the unimodal conditions. In the incongruent situation, participant preferentially categorized the affective expression based on the visual modality, demonstrating a visual dominance in emotional processing. However, when the reliability of the visual stimuli was diminished, participants categorized incongruent bimodal stimuli preferentially via the auditory modality. These results demonstrate that visual dominance in affect perception does not occur in a rigid manner, but follows flexible situation-dependent rules. In Experiment 2, we requested the participants to pay attention to only one sensory modality at a time in order to test the putative mandatory nature of multisensory affective interactions. We observed that even if they were asked to ignore concurrent sensory information, the irrelevant information significantly affected the processing of the target. This observation was especially true when the target modality was less reliable. Altogether, these findings indicate that the perception of emotion expressions is a robust multisensory situation which follows rules that have been previously observed in other perceptual domains.
The perception of emotions by ear and by eye.
DOI:10.1080/026999300378824 URL [本文引用: 3]
Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses
DOI:10.3758/BRM.41.4.1149 URL [本文引用: 2]
G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences
DOI:10.3758/BF03193146 URL [本文引用: 2]
Attentional bias for threat: Evidence for delayed disengagement from emotional faces
DOI:10.1080/02699930143000527 URL [本文引用: 1]
The competition between inhibition of return and emotional attention bias: Evidence from eye movements
返回抑制和情绪信息注意偏向的竞争:来自眼动的证据
Tracking the influence of reflexive attention on sensory and cognitive processing
The temporal dynamics of processing emotions from vocal, facial, and bodily expressions
DOI:10.1016/j.neuroimage.2011.06.035
PMID:21718792
[本文引用: 2]
Face-to-face communication works multimodally. Not only do we employ vocal and facial expressions; body language provides valuable information as well. Here we focused on multimodal perception of emotion expressions, monitoring the temporal unfolding of the interaction of different modalities in the electroencephalogram (EEG). In the auditory condition, participants listened to emotional interjections such as "ah", while they saw mute video clips containing emotional body language in the visual condition. In the audiovisual condition participants saw video clips with matching interjections. In all three conditions, the emotions "anger" and "fear", as well as non-emotional stimuli were used. The N100 amplitude was strongly reduced in the audiovisual compared to the auditory condition, suggesting a significant impact of visual information on early auditory processing. Furthermore, anger and fear expressions were distinct in the auditory but not the audiovisual condition. Complementing these event-related potential (ERP) findings, we report strong similarities in the alpha- and beta-band in the visual and the audiovisual conditions, suggesting a strong visual processing component in the perception of audiovisual stimuli. Overall, our results show an early interaction of modalities in emotional face-to-face communication using complex and highly natural stimuli.Copyright © 2011 Elsevier Inc. All rights reserved.
Do emotional faces affect inhibition of return? An ERP study
DOI:10.3389/fpsyg.2019.00001 URL [本文引用: 6]
Neural processing of emotion in multimodal settings
Inhibition of return
DOI:10.1016/S1364-6613(00)01452-2 URL [本文引用: 3]
The role of emotion in dynamic audiovisual integration of faces and voices
DOI:10.1093/scan/nsu105 URL [本文引用: 2]
Fear and anxiety: Animal models and human cognitive psychophysiology
The aim of this paper is to explicate what is special about emotional information processing, emphasizing the neural foundations that underlie the experience and expression of fear. A functional, anatomical model of defense behavior in animals is presented and applications are described in cognitive and physiological studies of human affect. It is proposed that unpleasant emotions depend on the activation of an evolutionarily primitive subcortical circuit, including the amygdala and the neural structures to which it projects. This motivational system mediates specific autonomic (e.g., heart rate change) and somatic reflexes (e.g., startle change) that originally promoted survival in dangerous conditions. These same response patterns are illustrated in humans, as they process objective, memorial, and media stimuli. Furthermore, it is shown how variations in the neural circuit and its outputs may separately characterize cue-specific fear (as in specific phobia) and more generalized anxiety. Finally, again emphasizing links between the animal and human data, we focus on special, attentional features of emotional processing: The automaticity of fear reactions, hyper-reactivity to minimal threat-cues, and evidence that the physiological responses in fear may be independent of slower, language-based appraisal processes.
Inhibition of return is unimpressed by emotional cues
DOI:10.1080/02699930701809451 URL [本文引用: 1]
The integration of facial expression and vocal emotion and its brain mechanism
DOI:10.3724/SP.J.1042.2019.01205 URL [本文引用: 1]
面孔表情和声音情绪信息整合加工的脑机制
Inhibition of return: Twenty years after
DOI:10.1080/02643290600588095 URL [本文引用: 1]
Timecourse of coactivation in bimodal divided attention
DOI:10.3758/BF03203025 URL [本文引用: 2]
Phobias and preparedness: The selective, automatic, and encapsulated nature of fear
We describe evidence for an evolved module for fear elicitation and fear learning with four primary characteristics. First, it is preferentially activated by stimuli related to survival threats in evolutionary history. Thus, fear-relevant stimuli lead to superior conditioning of aversive associations compared with fear-irrelevant stimuli. Second, the module is automatically activated by fear-relevant stimuli, meaning that fear activation occurs before conscious cognitive analysis of the stimulus can occur. Third, the fear module is relatively impenetrable to conscious cognitive control, and fear conditioning with fear-relevant stimuli can occur even with subliminal conditioned stimuli. Fourth, the amygdala seems to be the central brain area dedicated to the fear module. Finally, we propose that there are two levels of fear conditioning, with an emotional level that is relatively independent of the cognitive contingency level, each mediated by different brain areas.
Incongruence effects in crossmodal emotional integration
DOI:10.1016/j.neuroimage.2010.10.047 URL [本文引用: 1]
Increased inhibition following negative cues: A possible role for enhanced processing
DOI:S0010-9452(18)30418-0
PMID:30638583
[本文引用: 1]
Based on findings showing that attention is captured by aversive stimuli, previous studies have hypothesized that inhibition of return (IOR) is reduced at spatial locations previously occupied by threat cues. Yet evidence for this view is limited: Only a few studies have demonstrated a reduced degree of IOR following threat cues, while most have not found differences in IOR between aversive and neutral cues. In contrast to previous studies that used the spatial cuing paradigm and for the most part employed mild negative stimuli as cues, we examined the influence of highly aversive, colored and complex pictures of real life situations. As opposed to the stimuli used in previous studies, these pictures are thought to result in enhanced processing as well as in specific enhancement for threat pictures in comparison to neutral ones. Based on evidence indicating that enhanced processing of spatial cues results in increased IOR, we hypothesized that the negative picture cues employed in the present study would yield increased IOR. This hypothesis was confirmed in two experiments. We suggest that the enhancement of IOR following highly threatening cues may be related to efficient spatial orienting of attention in response to stimuli that are important from an evolutionary point of view. The results are discussed in the context of neurocognitive mechanisms that may underlie the modulation of IOR by emotional information.Copyright © 2018 Elsevier Ltd. All rights reserved.
Inhibition of return is modulated by negative stimuli: Evidence from subliminal perception
Attentional capture and trait anxiety: Evidence from inhibition of return
DOI:10.1016/j.janxdis.2009.03.002
PMID:19380211
[本文引用: 2]
Attentional biases regarding attentional capture by threat-related stimuli in anxious people were investigated by using a standard spatial cueing procedure suitable to measure inhibition of return (IOR). In two experiments, participants categorized the emotional valence of either emotional (positive and negative words) or non-emotional (neutral words in both experiments and sets of 'xxx' in Experiment 1) targets that were preceded by a peripheral non-predictive cue. The typical IOR effect (slower responses for words presented at previously cued locations) was observed for non-emotional and positive stimuli, with similar results being observed for both low and high trait anxiety groups. For negative stimuli, however, the high trait anxiety group did not show the IOR effect, while it was present in the low trait anxiety group. This general pattern of results suggests that, in individual with high trait anxiety, threatening stimuli can capture attention at the locations whether attentional capture is hindered by other cognitive effects such as IOR.
Reduced habituation to angry faces: Increased attentional capture as to override inhibition of return
DOI:10.1007/s00426-013-0493-9
PMID:23689476
[本文引用: 3]
The aim of this paper was to study whether real angry faces do capture attention to the extent of overcoming the inhibition of return (IOR) effect and whether the anxiety level of participants modulates this effect by stressing biases toward threatening stimuli. With this purpose, participants categorized the emotional valence of face targets in a standard spatial cueing procedure suitable to measure IOR. In Experiment 1, participants were selected according to their high vs. low-trait anxiety, whereas in Experiment 2 participants were induced a positive vs. anxiety mood state. The typical IOR effect was observed with neutral and happy face targets, which disappeared with angry face targets. Similar results were observed for all anxiety groups and in both experiments. The results indicate that IOR is overridden when the target is a biologically relevant angry face, as highly relevant targets should suffer less from habituation to attentional capture regardless of anxiety. We suggest that these data show that attentional capture is less likely to habituate for threatening information, so that no cost is measured in detecting new threatening information appearing at recently cued locations.
Components of visual orienting
The time-course of intermodal binding between seeing and hearing affective information
Intermodal binding between affective information that is seen as well as heard triggers a mandatory process of audiovisual integration. In order to track the time course of this audiovisual binding, event related brain potentials were recorded while subjects saw facial expression and concurrently heard auditory fragment. The results suggest that the combination of the two inputs is early in time (110 ms post-stimulus) and translates as a specific enhancement in amplitude of the auditory NI component. These findings are compatible with previous functional neuroimaging results of audiovisual speech showing strong audiovisual interactions in auditory cortex in the form of magnetic response amplifications, as well as with electrophysiological studies demonstrating early audiovisual interactions (before 200 ms post-stimulus). Moreover, our results show that the informational content present in the two modalities plays a crucial role in triggering the intermodal binding process.
Brain mechanisms for emotional influences on perception and attention: What is magic and what is not.
DOI:10.1016/j.biopsycho.2012.02.007
PMID:22373657
[本文引用: 1]
The rapid and efficient selection of emotionally salient or goal-relevant stimuli in the environment is crucial for flexible and adaptive behaviors. Converging data from neuroscience and psychology have accrued during the last decade to identify brain systems involved in emotion processing, selective attention, and their interaction, which together act to extract the emotional or motivational value of sensory events and respond appropriately. An important hub in these systems is the amygdala, which may not only monitor the emotional value of stimuli, but also readily project to several other areas and send feedback to sensory pathways (including striate and extrastriate visual cortex). This system generates saliency signals that modulate perceptual, motor, as well as memory processes, and thus in turn regulate behavior appropriately. Here, we review our current views on the function and properties of these brain systems, with an emphasis on their involvement in the rapid and/or preferential processing of threat-relevant stimuli. We suggest that emotion signals may enhance processing efficiency and competitive strength of emotionally significant events through gain control mechanisms similar to those of other (e.g. endogenous) attentional systems, but mediated by distinct neural mechanisms in amygdala and interconnected prefrontal areas. Alterations in these brain mechanisms might be associated with psychopathological conditions, such as anxiety or phobia. We conclude that attention selection and awareness are determined by multiple attention gain control systems that may operate in parallel and use different sensory cues but act on a common perceptual pathway.Copyright © 2012 Elsevier B.V. All rights reserved.
On the relationship between occipital cortex activity and inhibition of return
DOI:10.1111/psyp.2009.46.issue-6 URL [本文引用: 2]
Cortical expressions of inhibition of return
DOI:10.1016/j.brainres.2005.11.081 URL [本文引用: 1]
Inhibition of return: An information processing theory of its natures and significance
DOI:10.1016/j.cortex.2020.11.009
PMID:33360759
[本文引用: 1]
Inhibition of return (IOR) is an inhibitory aftereffect of visuospatial orienting, typically resulting in slower responses to targets presented in an area that has been recently attended. Since its discovery, myriad research has sought to explain the causes and effects underlying this phenomenon. Here, we briefly summarize the history of the phenomenon, and describe the early work supporting the functional significance of IOR as a foraging facilitator. We then shine a light on the discordance in the literature with respect to mechanism-in particular the lack of theoretical constructs that can consistently explain innumerable dissociations. We then describe three diagnostics (central arrow targets, locus of slack logic and the psychological refractory period, and performance in speed-accuracy space) used to support our theory that there are two forms of inhibition of return-the form which is manifest being contingent upon the activation state of the reflexive oculomotor system. The input form, which operates to decrease the salience of inputs, is generated when the reflexive oculomotor system is suppressed; the output form, which operates to bias responding, is generated when the reflexive oculomotor system is not suppressed. Then, we subject a published data set, wherein inhibitory effects had been generated while the reflexive oculomotor system was either active or suppressed, to diffusion modelling. As we hypothesized, based on the aforementioned theory, the effects of the two forms of IOR were best accounted for by different drift diffusion parameters. The paper ends with a variety of suggestions for further research.Copyright © 2020 Elsevier Ltd. All rights reserved.
What is inhibited in inhibition of return
DOI:10.1037/0096-1523.22.2.367 URL [本文引用: 1]
Effects of spatial cues on locating emotional targets
DOI:10.1080/13506280902787043 URL [本文引用: 2]
When do fearful faces override inhibition of return
DOI:10.1016/j.actpsy.2015.11.002 URL [本文引用: 5]
Multisensory integration: Current issues from the perspective of the single neuron
DOI:10.1038/nrn2331 URL [本文引用: 2]
Inhibition of return to social signals of fear
The present study examined whether inhibition of return (IOR) is modulated by the fear relevance of the cue. Experiment 1 found similar magnitude of IOR was produced by neutral and fear faces and luminance matched cues. To allow a more sensitive measure of endogenously directed attention, Experiment 2 removed a central reorienting cue and more precisely measured the time course of IOR. At stimulus onset asynchronies (SOAs) of 500, 1,000 and 1,500 ms, fear face and luminance matched cues resulted in similar IOR. These findings suggest that IOR is triggered by event onsets and disregards event value. Views of IOR as an adaptive "foraging facilitator," whereby attention is guided to promote optimal sampling of important environmental events, are discussed.(c) 2007 APA, all rights reserved.
Selective attention and multisensory integration: Multiple phases of effects on the evoked brain activity
We used event-related potentials (ERPs) to evaluate the role of attention in the integration of visual and auditory features of multisensory objects. This was done by contrasting the ERPs to multisensory stimuli (AV) to the sum of the ERPs to the corresponding auditory-only (A) and visual-only (V) stimuli [i.e., AV vs. (A + V)]. V, A, and VA stimuli were presented in random order to the left and right hemispaces. Subjects attended to a designated side to detect infrequent target stimuli in either modality there. The focus of this report is on the ERPs to the standard (i.e., nontarget) stimuli. We used rapid variable stimulus onset asynchronies (350-650 msec) to mitigate anticipatory activity and included "no-stim" trials to estimate and remove ERP overlap from residual anticipatory processes and from adjacent stimuli in the sequence. Spatial attention effects on the processing of the unisensory stimuli consisted of a modulation of visual P1 and N1 components (at 90-130 msec and 160-200 msec, respectively) and of the auditory N1 and processing negativity (100-200 msec). Attended versus unattended multisensory ERPs elicited a combination of these effects. Multisensory integration effects consisted of an initial frontal positivity around 100 msec that was larger for attended stimuli. This was followed by three phases of centro-medially distributed effects of integration and/or attention beginning at around 160 msec, and peaking at 190 (scalp positivity), 250 (negativity), and 300-500 msec (positivity) after stimulus onset. These integration effects were larger in amplitude for attended than for unattended stimuli, providing neural evidence that attention can modulate multisensory-integration processes at multiple stages.
Bimodal-divided attention attenuates visually induced inhibition of return with audiovisual targets
DOI:10.1007/s00221-019-05488-0 URL [本文引用: 7]
On the causes and effects of inhibition of return
DOI:10.3758/BF03208839 URL [本文引用: 1]
Inhibition of return for the discrimination of faces
DOI:10.3758/PP.70.2.279 URL [本文引用: 2]
A study on the neural mechanism of inhibition of return by the event-related potential in the Go/Nogo task
DOI:10.1016/j.biopsycho.2008.04.006
PMID:18524452
[本文引用: 1]
Inhibition of return (IOR) is a slowed response to a stimulus at recently cued locations when stimulus-onset asynchronies (SOAs) are longer than 250 ms. Using an uninformative peripheral cued Go/NoGo (commit/withdrawal response) task experiment, this study aimed to characterize the neural mechanism of IOR by studying not only the early event-related potentials (ERPs), P1 and N1, but also the late ERPs, Go/NoGo-N2 and P3. Scalp topographies and LORETA showed that the changes in P1 and N1, the cueing effects, were distributed mainly over the dorsal occipito-parietal areas, such as the bilateral middle occipital gyrus and the occipital portion of the cuneus. The changes in the late NoGo-N2 and P3 were distributed mainly over frontal-central areas, such as the right medial frontal gyrus. The NoGo-N2 was smaller and earlier in valid trials than in invalid trials, suggesting that the late component related to IOR was modulated by response preparation inhibition. The NoGo-P3 was larger and later in valid trials than in invalid trials, perhaps indicating that the control system (FEF) was free from an inhibitory marker in the cued locations. These data support a mechanism of IOR consisting of both sensory inhibition and response preparation inhibition.
The NimStim set of facial expressions: Judgments from untrained research participants
DOI:10.1016/j.psychres.2008.05.006
PMID:19564050
[本文引用: 1]
A set of face stimuli called the NimStim Set of Facial Expressions is described. The goal in creating this set was to provide facial expressions that untrained individuals, characteristic of research participants, would recognize. This set is large in number, multiracial, and available to the scientific community online. The results of psychometric evaluations of these stimuli are presented. The results lend empirical support for the validity and reliability of this set of facial expressions as determined by accurate identification of expressions and high intra-participant agreement across two testing sessions, respectively.
Unseen fearful faces promote amygdala guidance of attention
DOI:10.1093/scan/nss116 URL [本文引用: 1]
Testing the race model inequality: An algorithm and computer programs
In divided-attention tasks, responses are faster when two target stimuli are presented, and thus one is redundant, than when only a single target stimulus is presented. Raab (1962) suggested an account of this redundant-targets effect in terms of a race model in which the response to redundant target stimuli is initiated by the faster of two separate target detection processes. Such models make a prediction about the probability distributions of reaction times that is often called the race model inequality, and it is often of interest to test this prediction. In this article, we describe a precise algorithm that can be used to test the race model inequality and present MATLAB routines and a Pascal program that implement this algorithm.
Early multisensory interactions affect the competition among multiple visual objects
DOI:10.1016/j.neuroimage.2010.12.068 URL [本文引用: 1]
Visually induced inhibition of return affects the integration of auditory and visual information
DOI:10.1177/0301006616661934
PMID:27484341
[本文引用: 5]
Multisensory integration (MSI) and exogenous spatial attention can both speedup responses to perceptual events. Recently, it has been shown that audiovisual integration at exogenously attended locations is reduced relative to unattended locations. This effect was observed at short cue-target intervals (200-250 ms). At longer intervals, however, the initial benefits of exogenous shifts of spatial attention at the cued location are often replaced by response time (RT) costs (also known as Inhibition of Return, IOR). Given these opposing cueing effects at shorter versus longer intervals, we decided to investigate whether MSI would also be affected by IOR. Uninformative exogenous visual spatial cues were presented between 350 and 450 ms prior to the onset of auditory, visual, and audiovisual targets. As expected, IOR was observed for visual targets (invalid cue RT < valid cue RT). For auditory and audiovisual targets, neither IOR nor any spatial cueing effects were observed. The amount of relative multisensory response enhancement and race model inequality violation was larger for uncued as compared with cued locations indicating that IOR reduces MSI. The results are discussed in the context of changes in unisensory signal strength at cued as compared with uncued locations.
Dispositional fear, negative affectivity, and neuroimaging response to visually suppressed emotional faces
DOI:10.1016/j.neuroimage.2011.07.015 URL [本文引用: 1]
Influence of emotional faces for different location on inhibition of return
不同位置的情绪面孔加工对返回抑制的影响
Inhibition of return in static but not necessarily in dynamic search
DOI:10.3758/APP.72.1.76 URL [本文引用: 1]
Influence of inhibitory tagging (IT) on emotional and cognitive conflict processing: Evidence from event-related potentials
DOI:10.1016/j.neulet.2017.08.014 URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}