积极情绪对条件性恐惧泛化的抑制作用
1
2017
... 图形特征在知觉恐惧泛化研究中被广泛运用, 主要包括图形大小、颜色、形状等特征的变化.首先, 在图形大小方面, Lissek等(2008)曾用一大一小两个圆圈分别作为CS+和CS-, 8个大小处在CS+和CS-之间的圆圈作为GS测试被试的恐惧反应, 结果发现被试的眨眼惊跳反应随着GS与CS+大小的相似性增加而增强.随后, 研究者用该范式进一步确认, 广泛性焦虑障碍和惊恐障碍患者的恐惧泛化比正常被试更广(Lissek et al., 2010; Lissek, Kaczkurkin et al., 2014).近期, 使用该范式的国内研究发现, 状态焦虑会增加恐惧的泛化, 而积极情绪会减弱恐惧的泛化(徐亮等, 2016; 冯彪等, 2017). ...
状态焦虑对条件性恐惧泛化的影响
1
2016
... 图形特征在知觉恐惧泛化研究中被广泛运用, 主要包括图形大小、颜色、形状等特征的变化.首先, 在图形大小方面, Lissek等(2008)曾用一大一小两个圆圈分别作为CS+和CS-, 8个大小处在CS+和CS-之间的圆圈作为GS测试被试的恐惧反应, 结果发现被试的眨眼惊跳反应随着GS与CS+大小的相似性增加而增强.随后, 研究者用该范式进一步确认, 广泛性焦虑障碍和惊恐障碍患者的恐惧泛化比正常被试更广(Lissek et al., 2010; Lissek, Kaczkurkin et al., 2014).近期, 使用该范式的国内研究发现, 状态焦虑会增加恐惧的泛化, 而积极情绪会减弱恐惧的泛化(徐亮等, 2016; 冯彪等, 2017). ...
The impact of instructions on generalization of conditioned fear in humans
1
2015
... 图形形状特征变化通常结合颜色变化共同影响恐惧泛化.例如, Vervliet和Geens (2014)让被试对某一特定颜色的几何图形习得恐惧, CS-是含有CS+颜色或是形状特征的几何图形.泛化测试的结果表明, 被试对与CS-有相同特征的GS的恐惧反应较小.可见, 这项研究再次证明了CS-可以决定刺激泛化的模式.另外, Vervliet, Kindt, Vansteenwegen和Hermans (2010)发现口头指导语对恐惧泛化也有较明显的影响.他们将某一特定颜色的几何图形作为CS+, 在习得恐惧之前告知被试, 该几何图形的颜色或形状特征预示危险.结果发现被试对含有口头告知特征的GS更恐惧.还有研究者则认为该结果可能与注意和学习有关, 于是他们选择在习得之后呈现指导语, 而实验结果与前者一致(Ahmed & Lovibond 2015).可见, 认知过程在人类恐惧泛化中有着重要作用. ...
Generalization of contextual fear in humans
1
2015
... 在人类研究中, 实验者通常采用虚拟现实技术, 让被试进入两个不同的情景中, 其中一个场景匹配电击.泛化阶段则采用包含前两个情景事物的新异情景, 以此来评估被试的恐惧反应(Andreatta, Neueder, Glotzbach-Schoon, Mühlberger, & Pauli, 2017; Andreatta, Leombruni, Glotzbach-Schoon, Pauli, & Mühlberger, 2015).结果表明, 延迟2周之后呈现泛化情景比24小时之后呈现能引起更大恐惧反应(Andreatta et al., 2017).总的来说, 虚拟现实技术可以操纵情景特征的相似性, 使用虚拟现实技术或对治疗焦虑障碍有很大的促进作用.毕竟相对于暴露在真实的创伤情境下, 该技术显然具有安全和易操控的特点. ...
Effects of context preexposure and delay until anxiety retrieval on generalization of contextual anxiety
2
2017
... 在人类研究中, 实验者通常采用虚拟现实技术, 让被试进入两个不同的情景中, 其中一个场景匹配电击.泛化阶段则采用包含前两个情景事物的新异情景, 以此来评估被试的恐惧反应(Andreatta, Neueder, Glotzbach-Schoon, Mühlberger, & Pauli, 2017; Andreatta, Leombruni, Glotzbach-Schoon, Pauli, & Mühlberger, 2015).结果表明, 延迟2周之后呈现泛化情景比24小时之后呈现能引起更大恐惧反应(Andreatta et al., 2017).总的来说, 虚拟现实技术可以操纵情景特征的相似性, 使用虚拟现实技术或对治疗焦虑障碍有很大的促进作用.毕竟相对于暴露在真实的创伤情境下, 该技术显然具有安全和易操控的特点. ...
... ).结果表明, 延迟2周之后呈现泛化情景比24小时之后呈现能引起更大恐惧反应(Andreatta et al., 2017).总的来说, 虚拟现实技术可以操纵情景特征的相似性, 使用虚拟现实技术或对治疗焦虑障碍有很大的促进作用.毕竟相对于暴露在真实的创伤情境下, 该技术显然具有安全和易操控的特点. ...
BDNF in the dentate gyrus is required for consolidation of “pattern-separated” memories
1
2013
... 情景的恐惧泛化需要对整个情景内的信息进行辨别, 这些信息通常包括空间、嗅觉、听觉和视觉信息, 而不是单一的物体特征的变化(Jasnow, Lynch, Gilman, & Riccio, 2017).首先, 在动物研究方面, 有研究者训练啮齿类动物对某一情景(CXT+)习得恐惧, 而在另外一个新异的情景(CXT-)下始终不匹配恐惧刺激, 紧接着在一个和CXT+相似的情景(G-CXT)中进行恐惧泛化测试.这些情景任务包括辨别触摸屏中物体与屏幕的距离(Mcallister, Saksida, & Bussey, 2013; Talpos, McTighe, Dias, Saksida, & Bussey, 2010)、辨别新异刺激和熟悉刺激的位置(Bekinschtein et al., 2013)、在不同形状的空间里习得恐惧(Mchugh et al., 2007)等.动物研究表明, 恐惧习得和重现刺激情景(G-CXT)之间的时间间隔越长, 恐惧泛化程度越大(Biedenkapp & Rudy, 2007).而在习得任务之前预先暴露情景则会减弱恐惧的泛化.此外, 研究者通过运用情景恐惧泛化范式, 发现啮齿动物恐惧泛化存在着性别差异, 具体表现为雌性比雄性更容易发生泛化(Keiser et al., 2017), 且泛化速度更快(Lynch, Cullen, Jasnow, & Riccio, 2013).预先暴露情景增加了雄性老鼠在习得阶段的恐惧反应, 但却减少了雌性老鼠在泛化的阶段恐惧反应(Keiser et al., 2017). ...
From bad to worse: Symbolic equivalence and opposition in fear generalisation
1
2015
... 从研究内容上来说, 实验室所采用的刺激都是单一的感觉线索, 如图形大小、颜色、声调等, 而在现实生活中的恐惧习得却包含了大量的复杂刺激, 这些刺激通常是多维的, 比如战场、猛兽等.可以说, 基于知觉的恐惧泛化研究低估了现实生活中恐惧泛化的复杂性.另外, 人们常常会根据经验, 去归纳、推断情绪事件的未知属性以及因果联系(Dunsmoor & Murphy, 2015).例如, 一个人对针头产生了恐惧经验, 他就可能对包括医院、护士等在概念上相关的事物产生害怕心理.因此, 基于知觉的恐惧泛化及其相关理论并不能完全解释人类恐惧泛化的现象.例如, Dunsmoor和Murphy (2014)探索了概念的典型性对恐惧泛化的影响, 结果发现对典型成员习得的恐惧会泛化到非典型成员身上, 但非典型成员习得的恐惧则不能泛化到典型成员身上.虽然已有少量研究注意到了如语义、类别、符号等基于高级认知过程的恐惧泛化现象(Bennett, Hermans, Dymond, Vervoort, & Baeyens, 2015; Dunsmoor, White, & LaBar, 2011; Dymond, Schlund, Roche, & Whelan, 2014), 但这方面的实证研究仍然很薄弱, 尚有许多实际生活中的问题不能得到合理解释, 如一个人被金毛犬咬过之后, 是会对所有的狗产生恐惧还是只对金毛犬产生恐惧?又如何解释上述对“针头”产生恐惧并泛化到“医院”上的现象?因此未来的研究应关注高级认知过程与恐惧泛化的交互作用, 以期完善恐惧泛化的理论基础, 并对相关临床疾病的治疗提供帮助. ...
Neurobiological substrates of dread
1
2006
... 脑岛和海马、杏仁核、水管周灰质(periaqueductal gray )共同控制自主神经系统, 它们在加工恐惧情绪时被激活(Saper, 1982).前人研究表明, 当个体面对不确定的情景及对恐惧刺激进行预期时, 脑岛的响应随之增强(Berns et al., 2006; Dunsmoor, Bandettini, & Knight, 2007).一般认为, 前脑岛是躯体感觉和内脏感觉的结合点, 与ACC、杏仁核、辅助运动区(supplementary motor area, SMA)等区域一起引发与恐惧相关的行为.Dunsmoor, Prince等(2011)首次用功能成像技术结合情绪面孔感知维度对人类恐惧泛化进行研究, 并证明了在条件性恐惧的习得和表达中, 当GS接近CS时, 脑岛、尾状核(caudate)、丘脑的激活程度普遍增加.例如Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013a)在用几何图形测试人类恐惧泛化的过程中, 发现在接近CS时, 脑岛、尾状核、ACC、SMA的响应会随着刺激相似性的增加而增强.随后, Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013b)用相同的范式对广泛性焦虑障碍患者进行恐惧泛化测试, 发现前脑岛(anterior insular, AI)与ACC、杏仁核、SMA连接增强.同时, Lissek, Bradford 等(2014)用圆圈刺激发现在恐惧泛化任务中, 正常被试脑岛激活增强.此后, Onat和Büchel (2015)用面孔范式发现脑岛对CS+最敏感, 颞下皮层(inferotemporal cortex, ITC)对CS+和CS-中间的模糊刺激比较敏感, 这表明前脑岛皮层编码“危险识别”过程, 颞下皮层则对“模糊不清”的信息进行编码.可见, 脑岛在知觉的恐惧泛化过程中参与恐惧的表达和刺激的辨别. ...
Context preexposure prevents forgetting of a contextual fear memory: Implication for regional changes in brain activation patterns associated with recent and remote memory tests
1
2007
... 情景的恐惧泛化需要对整个情景内的信息进行辨别, 这些信息通常包括空间、嗅觉、听觉和视觉信息, 而不是单一的物体特征的变化(Jasnow, Lynch, Gilman, & Riccio, 2017).首先, 在动物研究方面, 有研究者训练啮齿类动物对某一情景(CXT+)习得恐惧, 而在另外一个新异的情景(CXT-)下始终不匹配恐惧刺激, 紧接着在一个和CXT+相似的情景(G-CXT)中进行恐惧泛化测试.这些情景任务包括辨别触摸屏中物体与屏幕的距离(Mcallister, Saksida, & Bussey, 2013; Talpos, McTighe, Dias, Saksida, & Bussey, 2010)、辨别新异刺激和熟悉刺激的位置(Bekinschtein et al., 2013)、在不同形状的空间里习得恐惧(Mchugh et al., 2007)等.动物研究表明, 恐惧习得和重现刺激情景(G-CXT)之间的时间间隔越长, 恐惧泛化程度越大(Biedenkapp & Rudy, 2007).而在习得任务之前预先暴露情景则会减弱恐惧的泛化.此外, 研究者通过运用情景恐惧泛化范式, 发现啮齿动物恐惧泛化存在着性别差异, 具体表现为雌性比雄性更容易发生泛化(Keiser et al., 2017), 且泛化速度更快(Lynch, Cullen, Jasnow, & Riccio, 2013).预先暴露情景增加了雄性老鼠在习得阶段的恐惧反应, 但却减少了雌性老鼠在泛化的阶段恐惧反应(Keiser et al., 2017). ...
A modern learning theory perspective on the etiology of panic disorder
1
2001
... 然而, 过度的恐惧泛化则会对个体生存产生不利影响.一方面, 过度的恐惧泛化被认为是广泛性焦虑障碍(generalized anxiety disorder, GAD)、创伤后应激障碍(post-traumatic stress disorder, PTSD)、惊恐障碍(panic disorder, PD)、特殊恐惧症(Specific phobia)等焦虑障碍的重要致病因素之一(Onat & Büchel, 2015), 如个体遭受严重车祸后, 在面对与车祸相关的线索或者情景(如红绿灯路口、鸣笛声等)时则会表现出恐惧情绪及回避反应; 另一方面, 恐惧泛化不仅仅限于外在刺激, 如果个体在经历创伤性事件时有心跳加速的症状, 那么与此相联系的其他内在感觉, 如胸闷、头昏的症状也可能引发恐惧(Bouton, Mineka, & Barlow, 2001).由此可见, 过度的恐惧泛化将会导致个体过度焦虑、社会功能受损和生活质量下降等诸多问题, 从而给患者的正常生活造成巨大困扰.因此, 恐惧泛化研究对克服焦虑障碍具有重要的临床意义. ...
Effect of post-retrieval propranolol on psychophysiologic responding during subsequent script- driven traumatic imagery in post-traumatic stress disorder
1
2008
... 此外, 去甲肾上腺素(norepinephrine, NE)也已成功应用于PTSD的治疗与防治(Brunet et al., 2008; Pervanidou & Chrousos, 2010).动物实验表明, 去甲肾上腺素可能通过调节小鼠内侧前额叶皮层(mPFC)参与调节恐惧的回路, 从而对消退恐惧障碍起到改善作用(Fitzgerald, Giustino, Seemann, & Maren, 2015). ...
Attending to and remembering tactile stimuli: A review of brain imaging data and single-neuron responses
1
2000
... 从研究方法上来说, 刺激的不同设置方式是否会影响被试的辨别力?首先, 在对GS恐惧程度进行评估时, 刺激单独呈现, 并被较长的时间间隔分开, 这就减弱了刺激间直接的对比.此外, 不同的时间间隔将会触发不同的记忆过程, 因为随着时间间隔的增长, 被试的行为反应会相应减弱(Burton & Sinclair, 2000).其次, 被试的情绪状态也会影响实验结果.在单纯的知觉辨别任务中, 没有呈现厌恶刺激时被试的情绪是相对中性的.但在恐惧泛化任务中, 恐惧的情绪状态会影响被试的决策并减少知觉的准确性, 从而影响被试对GS和CS的辨别(Stefanucci & Storbeck, 2009).由此可见, 个体在恐惧状态和正常状态下的辨别能力是存在差异的.再次, 刺激梯度通常是人为操纵, 而非基于被试的知觉, 而个体的感知阈限不同, 对刺激的辨别度也存在差异, 如Resnik, Sobel和Paz (2011)用纯音(1/2 kHz)分别作为CS+和CS-, 被试却经常把CS+周围的刺激认作CS+.然而, 只有极少数研究用差别阈限法关注被试对材料的辨别度(Holt et al., 2014).所以未来的研究应该注意区分被试对刺激的辨别力, 采用差别阈限法对实验材料进行设置以避免因混淆辨别力不佳而造成的实验结果误差. ...
Circuit-wide structural and functional measures predict ventromedial prefrontal cortex fear generalization: Implications for generalized anxiety disorder
1
2014
... 内侧前额叶皮层(medial prefrontal cortex, mPFC)是恐惧神经环路的关键脑区, 在恐惧消退过程中起着抑制恐惧的作用(Milad et al., 2007; Orsini & Maren, 2012).近来研究表明, mPFC与恐惧泛化过程也有密切的联系.Cullen, Gilman, Winiecki, Riccio和Jasnow (2015)发现, 情景泛化记忆的表达与前扣带皮层(anterior cingulate cortex, ACC)、下边缘皮层(infralimbic cortex, IL)、前边缘皮层(prelimbic cortex, PL)这些前额叶皮层脑区的活性有关, 其中ACC与海马CA1区的连接抑制将减弱恐惧的泛化.另外, 与脑岛等脑区相反的是, vmPFC在接近CS-时激活更强(Greenberg et al., 2013a, 2013b; Lissek, Brandford et al., 2014), 在安全线索条件下GAD患者的vmPFC激活较正常人弱(Greenberg et al., 2013b).此外, vmPFC皮质厚度和恐惧泛化梯度有关, vmPFC皮质的减少可能是导致GAD患者过度泛化的原因之一(Cha et al, 2014).Xu等(2012)观察到mPFC处突触传递的毁损将导致动物情景恐惧记忆的过度泛化.他们认为mPFC、海马以及连接核(nucleus reuniens, NR) (丘脑中线核群的一部分)共同控制着恐惧记忆的泛化.具体而言, 由于mPFC是参与动机及情绪状态的关键脑区, 因此mPFC-NR路径将记忆的动机以及情绪效价信息传递至海马进行特征的重叠编码, 海马又将编码后的信息返回至mPFC进而对泛化记忆进行调节(Xu & Südhof, 2013).另有研究表明, 老鼠对声音习得的恐惧反应和mPFC处胆碱能系统(nitrergic system) (神经元轴突分泌的一类神经递质)的激活水平能够预测泛化的程度, 表现为胆碱能系统激活程度越高, 泛化程度越高(Saul’skaya & Sudorgina, 2016).此外, Likhtik, Stujenske, Topiwala, Harris和Gordon (2014)认为内侧前额叶(mPFC)以及基底外侧核(BLA)连接的增强预示着内侧前额叶可能会影响基底外侧核的活动, 并通过抑制杏仁核的活动来减弱恐惧反应.相反的, mPFC-BLA连接的减弱则会导致对杏仁核的抑制失败, 从而增强恐惧反应.可见, mPFC在知觉恐惧泛化过程中参与调节和抑制恐惧. ...
Encoding of conditioned fear in central amygdala inhibitory circuits
1
2010
... 杏仁核是恐惧学习和恐惧泛化的关键脑区, 是CS与US的汇聚点并启动由威胁引起的条件反应.运用情景辨别任务, Ciocchi等(2010)发现激活中央杏仁核外侧核(lateral part of central amygdala, CeL)神经元的活动, 或者抑制中央杏仁核内侧核(medial part of central amygdala, CeM)神经元的活动都会增加老鼠对CS-的恐惧反应.这说明杏仁核中央核团(central amygdala, CeA)的神经活性对恐惧泛化具有一定的调节作用.Lopresto, Schipper和Homberg (2016)认为探索杏仁核在恐惧泛化中的作用即是把杏仁核看作神经回路的节点, 该节点连接着海马和大脑皮层.在情景任务中, 已有研究表明恐惧情景引发老鼠和人类的杏仁核以及海马的响应比中性情景更强(Maren, Phan, & Liberzon, 2013; Marschner, Kalisch, Vervliet, Vansteenwegen, & Buchel, 2008).这说明, 杏仁核以及海马可能是调节情景恐惧泛化的关键脑区之一.此后, Resnik和Paz (2015)用纯音作为刺激材料考察了猴子恐惧泛化的神经机制及其行为反应, 发现基底外侧核(basolateral complex of the amygdala, BLA)处神经元的特异变化减弱了泛化阶段的辨别学习, 这种杏仁核部位的轻微改变影响了前额叶皮层以及海马, 并破坏了泛化的平衡, 从而导致了更广的泛化.另外, 内侧前额叶(mPFC)-BLA通路的增强可以促进恐惧消退, 这种自上而下连接不良的传递, 也会导致更广的泛化(Jasnow et al., 2017; Likhtik & Paz, 2015).由此可见, 杏仁核在知觉恐惧泛化回路中有着重要的作用, 是其他脑区与感知觉信息的连接点, 控制着恐惧反应的表达. ...
Activity of the anterior cingulate cortex and ventral hippocampus underlie increases in contextual fear generalization
1
2015
... 内侧前额叶皮层(medial prefrontal cortex, mPFC)是恐惧神经环路的关键脑区, 在恐惧消退过程中起着抑制恐惧的作用(Milad et al., 2007; Orsini & Maren, 2012).近来研究表明, mPFC与恐惧泛化过程也有密切的联系.Cullen, Gilman, Winiecki, Riccio和Jasnow (2015)发现, 情景泛化记忆的表达与前扣带皮层(anterior cingulate cortex, ACC)、下边缘皮层(infralimbic cortex, IL)、前边缘皮层(prelimbic cortex, PL)这些前额叶皮层脑区的活性有关, 其中ACC与海马CA1区的连接抑制将减弱恐惧的泛化.另外, 与脑岛等脑区相反的是, vmPFC在接近CS-时激活更强(Greenberg et al., 2013a, 2013b; Lissek, Brandford et al., 2014), 在安全线索条件下GAD患者的vmPFC激活较正常人弱(Greenberg et al., 2013b).此外, vmPFC皮质厚度和恐惧泛化梯度有关, vmPFC皮质的减少可能是导致GAD患者过度泛化的原因之一(Cha et al, 2014).Xu等(2012)观察到mPFC处突触传递的毁损将导致动物情景恐惧记忆的过度泛化.他们认为mPFC、海马以及连接核(nucleus reuniens, NR) (丘脑中线核群的一部分)共同控制着恐惧记忆的泛化.具体而言, 由于mPFC是参与动机及情绪状态的关键脑区, 因此mPFC-NR路径将记忆的动机以及情绪效价信息传递至海马进行特征的重叠编码, 海马又将编码后的信息返回至mPFC进而对泛化记忆进行调节(Xu & Südhof, 2013).另有研究表明, 老鼠对声音习得的恐惧反应和mPFC处胆碱能系统(nitrergic system) (神经元轴突分泌的一类神经递质)的激活水平能够预测泛化的程度, 表现为胆碱能系统激活程度越高, 泛化程度越高(Saul’skaya & Sudorgina, 2016).此外, Likhtik, Stujenske, Topiwala, Harris和Gordon (2014)认为内侧前额叶(mPFC)以及基底外侧核(BLA)连接的增强预示着内侧前额叶可能会影响基底外侧核的活动, 并通过抑制杏仁核的活动来减弱恐惧反应.相反的, mPFC-BLA连接的减弱则会导致对杏仁核的抑制失败, 从而增强恐惧反应.可见, mPFC在知觉恐惧泛化过程中参与调节和抑制恐惧. ...
Modulation of aversive memory by adult hippocampal neurogenesis
1
2017
... 海马中 “模式分离(pattern separation)”和“模式完成(pattern completion)”负责对刺激的属性特征与已存储的信息进行匹配(Gluck & Myers, 1993).“模式分离”由海马的齿状回执行(dentate gyrus, DG), 是一种把相似信息区分开的神经活动; 而“模式完成”主要发生在海马CA3区, 它描述了相反的加工过程, 如果刺激的区别较小, “模式完成”则会忽略刺激间的区别(Drew & Huckleberry, 2017; Gilbert, Kesner, & Lee, 2001; Treves & Rolls, 1992).在知觉泛化过程中, 当GS和CS相似性较高时, 海马启动“模式完成”使CS的神经表征激活, 从而进一步引发被试的恐惧反应; 而当GS和CS相似性较低时, 海马则启动“模式分离”激活腹内测前额叶从而降低杏仁核的活动, 达到抑制恐惧反应的效果(Lissek, 2012; Lissek, Bradford et al., 2014).例如, 研究发现在啮齿动物的情景辨别恐惧泛化任务中, 海马CA3区的神经抑制会促进啮齿动物在恐惧和中性情景下的“模式完成”过程, 从而增加恐惧的泛化; 反过来, 齿状回处的神经兴奋则会促进“模式分离”过程, 以抑制恐惧泛化(Sahay et al., 2011; Niibori et al., 2012).可见, 海马在知觉恐惧泛化的过程中负责刺激的辨别加工. ...
Impact of continuous versus intermittent CS-UCS pairing on human brain activation during Pavlovian fear conditioning
1
2007
... 脑岛和海马、杏仁核、水管周灰质(periaqueductal gray )共同控制自主神经系统, 它们在加工恐惧情绪时被激活(Saper, 1982).前人研究表明, 当个体面对不确定的情景及对恐惧刺激进行预期时, 脑岛的响应随之增强(Berns et al., 2006; Dunsmoor, Bandettini, & Knight, 2007).一般认为, 前脑岛是躯体感觉和内脏感觉的结合点, 与ACC、杏仁核、辅助运动区(supplementary motor area, SMA)等区域一起引发与恐惧相关的行为.Dunsmoor, Prince等(2011)首次用功能成像技术结合情绪面孔感知维度对人类恐惧泛化进行研究, 并证明了在条件性恐惧的习得和表达中, 当GS接近CS时, 脑岛、尾状核(caudate)、丘脑的激活程度普遍增加.例如Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013a)在用几何图形测试人类恐惧泛化的过程中, 发现在接近CS时, 脑岛、尾状核、ACC、SMA的响应会随着刺激相似性的增加而增强.随后, Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013b)用相同的范式对广泛性焦虑障碍患者进行恐惧泛化测试, 发现前脑岛(anterior insular, AI)与ACC、杏仁核、SMA连接增强.同时, Lissek, Bradford 等(2014)用圆圈刺激发现在恐惧泛化任务中, 正常被试脑岛激活增强.此后, Onat和Büchel (2015)用面孔范式发现脑岛对CS+最敏感, 颞下皮层(inferotemporal cortex, ITC)对CS+和CS-中间的模糊刺激比较敏感, 这表明前脑岛皮层编码“危险识别”过程, 颞下皮层则对“模糊不清”的信息进行编码.可见, 脑岛在知觉的恐惧泛化过程中参与恐惧的表达和刺激的辨别. ...
Threat intensity widens fear generalization gradients
1
2017
... 听觉是仅次于视觉的重要感觉通道, 也是动物和人类感知环境、逃避危险的线索之一.在恐惧泛化的研究中, 研究者们根据频率把声音划分成等距且可辨别的几段音频, 并与电击匹配, 来研究听觉恐惧泛化的机制.动物研究中对恐惧泛化脑机制的探索通常采用声音辨别任务(Ghosh & Chattarji, 2015; Resnik & Paz, 2015).Hovland等(1937)首次将该方法用以研究人类的恐惧泛化, 实验采用4个振幅相同、声音频率等距且可辨别的纯音作为材料, 结果发现被试对从未与US匹配的其他三个纯音也会产生恐惧反应.此外, 研究者用声音结合不同强度的电击, 探讨不同强度电击对恐惧泛化的影响, 结果表明, 不同强度的电击对被试习得恐惧并没有产生影响, 但高强度的电击却可以增加泛化的范围(Dunsmoor, Kroes, Braren, & Phelps, 2017). ...
Brain activity associated with omission of an aversive event reveals the effects of fear learning and generalization
1
2012
... 第二类研究是同一个体的不同程度恐惧的面孔, 其中中等恐惧的面孔作为CS+, 低恐惧的面孔作为CS-.结果发现, 被试对高恐惧面孔而不是中等恐惧面孔表现出更大的恐惧反应.这表明, 情绪强度比知觉的相似程度更能驱动恐惧的泛化(Dunsmoor, Prince et al., 2011; Dunsmoor & LaBar, 2012). ...
Effects of discrimination training on fear generalization gradients and perceptual classification in humans
1
2013
... 图形颜色特征变化也很常见.与大小变化相似, 研究者通常采用两个颜色差别明显的圆圈分别作为CS+和CS-, 再用颜色处在之间变化的圆圈测试被试的恐惧反应.例如Lommen, Engelhard和van den Hout (2010)采用白色(CS+)、黑色(CS-)和灰色(GS)圆圈作为实验材料, 发现神经质特质人群的恐惧泛化存在着显著差异, 表现为高神经质组在面对模棱两可的刺激时表现出比低神经质组更多的恐惧反应.Dunsmoor和LaBar (2013)通过操纵CS-的颜色来探讨CS-特征对恐惧泛化的影响, 即用蓝绿色圆圈作为CS+, 绿色或蓝色圆圈作为CS-, 处在CS+和CS-之间的变化的圆圈测试恐惧泛化反应, 结果发现被试对远离CS-的刺激产生了泛化. ...
Categories, concepts, and conditioning: How humans generalize fear
1
2015
... 从研究内容上来说, 实验室所采用的刺激都是单一的感觉线索, 如图形大小、颜色、声调等, 而在现实生活中的恐惧习得却包含了大量的复杂刺激, 这些刺激通常是多维的, 比如战场、猛兽等.可以说, 基于知觉的恐惧泛化研究低估了现实生活中恐惧泛化的复杂性.另外, 人们常常会根据经验, 去归纳、推断情绪事件的未知属性以及因果联系(Dunsmoor & Murphy, 2015).例如, 一个人对针头产生了恐惧经验, 他就可能对包括医院、护士等在概念上相关的事物产生害怕心理.因此, 基于知觉的恐惧泛化及其相关理论并不能完全解释人类恐惧泛化的现象.例如, Dunsmoor和Murphy (2014)探索了概念的典型性对恐惧泛化的影响, 结果发现对典型成员习得的恐惧会泛化到非典型成员身上, 但非典型成员习得的恐惧则不能泛化到典型成员身上.虽然已有少量研究注意到了如语义、类别、符号等基于高级认知过程的恐惧泛化现象(Bennett, Hermans, Dymond, Vervoort, & Baeyens, 2015; Dunsmoor, White, & LaBar, 2011; Dymond, Schlund, Roche, & Whelan, 2014), 但这方面的实证研究仍然很薄弱, 尚有许多实际生活中的问题不能得到合理解释, 如一个人被金毛犬咬过之后, 是会对所有的狗产生恐惧还是只对金毛犬产生恐惧?又如何解释上述对“针头”产生恐惧并泛化到“医院”上的现象?因此未来的研究应关注高级认知过程与恐惧泛化的交互作用, 以期完善恐惧泛化的理论基础, 并对相关临床疾病的治疗提供帮助. ...
Stimulus typicality determines how broadly fear is generalized
1
2014
... 从研究内容上来说, 实验室所采用的刺激都是单一的感觉线索, 如图形大小、颜色、声调等, 而在现实生活中的恐惧习得却包含了大量的复杂刺激, 这些刺激通常是多维的, 比如战场、猛兽等.可以说, 基于知觉的恐惧泛化研究低估了现实生活中恐惧泛化的复杂性.另外, 人们常常会根据经验, 去归纳、推断情绪事件的未知属性以及因果联系(Dunsmoor & Murphy, 2015).例如, 一个人对针头产生了恐惧经验, 他就可能对包括医院、护士等在概念上相关的事物产生害怕心理.因此, 基于知觉的恐惧泛化及其相关理论并不能完全解释人类恐惧泛化的现象.例如, Dunsmoor和Murphy (2014)探索了概念的典型性对恐惧泛化的影响, 结果发现对典型成员习得的恐惧会泛化到非典型成员身上, 但非典型成员习得的恐惧则不能泛化到典型成员身上.虽然已有少量研究注意到了如语义、类别、符号等基于高级认知过程的恐惧泛化现象(Bennett, Hermans, Dymond, Vervoort, & Baeyens, 2015; Dunsmoor, White, & LaBar, 2011; Dymond, Schlund, Roche, & Whelan, 2014), 但这方面的实证研究仍然很薄弱, 尚有许多实际生活中的问题不能得到合理解释, 如一个人被金毛犬咬过之后, 是会对所有的狗产生恐惧还是只对金毛犬产生恐惧?又如何解释上述对“针头”产生恐惧并泛化到“医院”上的现象?因此未来的研究应关注高级认知过程与恐惧泛化的交互作用, 以期完善恐惧泛化的理论基础, 并对相关临床疾病的治疗提供帮助. ...
Fear generalization and anxiety: Behavioral and neural mechanisms
1
2015
... 为了避免受到既有经验的影响, 实验室研究通常采用简单的感觉信息(如纯音、简单几何图形、面孔)作为实验材料, 同时结合电击、恐惧的面孔、厌恶的气味等负性刺激进行基于知觉的恐惧泛化测试(Dunsmoor & Paz, 2015; Dymond et al., 2015).根据知觉信息的不同维度, 可分为视觉、听觉、情景三方面, 以下将逐一论述. ...
Neurobehavioral mechanisms of human fear generalization
4
2011
... 恐惧是一种对环境的适应性机制, 当危险发生时, 恐惧使得机体启动防御机制并对危险刺激进行反应(Dymond, Dunsmoor, Vervliet, Roche, & Hermans, 2015; Hoppenbrouwers, Bulten, & Brazil, 2016; LeDoux, 2000).人类的恐惧反应多是后天习得, 例如, 在华生的条件反射实验中, 小阿尔伯特原本并不惧怕白鼠, 然而当白鼠与巨大的金属响声(危险刺激)同时出现后, 小阿尔伯特便对白鼠产生了恐惧反应, 此后当他再见到小白兔、白色绒毛玩具、甚至老人的白胡须等相关事物时, 他都会感到害怕(Watson & Rayner, 1920).这种不仅对特定的刺激(白鼠)产生恐惧, 而且对与特定刺激相关的事物同样会产生恐惧的现象即是“恐惧泛化” (Dunsmoor, Prince, Murty, Kragel, & LaBar, 2011; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014; Onat & Büchel, 2015).一般来说, 适当的恐惧泛化对个体生存具有积极意义, 因为在漫长的进化过程中, 恐惧泛化能够促使人类躲避各种潜在威胁, 有利于个体生存繁衍; 而在日常生活中, 恐惧泛化也可增强个体的警觉性, 从而有效并快速地对危险环境做出预测和应对(Steimer, 2002). ...
... 第二类研究是同一个体的不同程度恐惧的面孔, 其中中等恐惧的面孔作为CS+, 低恐惧的面孔作为CS-.结果发现, 被试对高恐惧面孔而不是中等恐惧面孔表现出更大的恐惧反应.这表明, 情绪强度比知觉的相似程度更能驱动恐惧的泛化(Dunsmoor, Prince et al., 2011; Dunsmoor & LaBar, 2012). ...
... 脑岛和海马、杏仁核、水管周灰质(periaqueductal gray )共同控制自主神经系统, 它们在加工恐惧情绪时被激活(Saper, 1982).前人研究表明, 当个体面对不确定的情景及对恐惧刺激进行预期时, 脑岛的响应随之增强(Berns et al., 2006; Dunsmoor, Bandettini, & Knight, 2007).一般认为, 前脑岛是躯体感觉和内脏感觉的结合点, 与ACC、杏仁核、辅助运动区(supplementary motor area, SMA)等区域一起引发与恐惧相关的行为.Dunsmoor, Prince等(2011)首次用功能成像技术结合情绪面孔感知维度对人类恐惧泛化进行研究, 并证明了在条件性恐惧的习得和表达中, 当GS接近CS时, 脑岛、尾状核(caudate)、丘脑的激活程度普遍增加.例如Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013a)在用几何图形测试人类恐惧泛化的过程中, 发现在接近CS时, 脑岛、尾状核、ACC、SMA的响应会随着刺激相似性的增加而增强.随后, Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013b)用相同的范式对广泛性焦虑障碍患者进行恐惧泛化测试, 发现前脑岛(anterior insular, AI)与ACC、杏仁核、SMA连接增强.同时, Lissek, Bradford 等(2014)用圆圈刺激发现在恐惧泛化任务中, 正常被试脑岛激活增强.此后, Onat和Büchel (2015)用面孔范式发现脑岛对CS+最敏感, 颞下皮层(inferotemporal cortex, ITC)对CS+和CS-中间的模糊刺激比较敏感, 这表明前脑岛皮层编码“危险识别”过程, 颞下皮层则对“模糊不清”的信息进行编码.可见, 脑岛在知觉的恐惧泛化过程中参与恐惧的表达和刺激的辨别. ...
... Laufer等(2016)用声音刺激发现, 相比于正常被试, GAD被试的dACC、尾状核、初级听觉皮层(primary auditory cortex)激活更强; 而正常被试的vmPFC和腹侧前扣带回 (ventral ACC) 则强于GAD被试.另外GAD被试在习得阶段dACC响应与负性刺激的泛化相关; 而尾状核(putamen)的响应与正性刺激的泛化相关.这表明在恐惧学习阶段的脑区响应对恐惧泛化阶段有一定预测作用.Dunsmoor, Prince等(2011)用面孔刺激发现泛化阶段杏仁核与负责面孔识别的脑区——梭状回(fusiform gyrus, FFG)的连接增强.由此可见, 杏仁核与初级感觉皮层的连接通过提高恐惧相关联的感觉信息来促进恐惧的表达. ...
Conceptual similarity promotes generalization of higher order fear learning
1
2011
... 从研究内容上来说, 实验室所采用的刺激都是单一的感觉线索, 如图形大小、颜色、声调等, 而在现实生活中的恐惧习得却包含了大量的复杂刺激, 这些刺激通常是多维的, 比如战场、猛兽等.可以说, 基于知觉的恐惧泛化研究低估了现实生活中恐惧泛化的复杂性.另外, 人们常常会根据经验, 去归纳、推断情绪事件的未知属性以及因果联系(Dunsmoor & Murphy, 2015).例如, 一个人对针头产生了恐惧经验, 他就可能对包括医院、护士等在概念上相关的事物产生害怕心理.因此, 基于知觉的恐惧泛化及其相关理论并不能完全解释人类恐惧泛化的现象.例如, Dunsmoor和Murphy (2014)探索了概念的典型性对恐惧泛化的影响, 结果发现对典型成员习得的恐惧会泛化到非典型成员身上, 但非典型成员习得的恐惧则不能泛化到典型成员身上.虽然已有少量研究注意到了如语义、类别、符号等基于高级认知过程的恐惧泛化现象(Bennett, Hermans, Dymond, Vervoort, & Baeyens, 2015; Dunsmoor, White, & LaBar, 2011; Dymond, Schlund, Roche, & Whelan, 2014), 但这方面的实证研究仍然很薄弱, 尚有许多实际生活中的问题不能得到合理解释, 如一个人被金毛犬咬过之后, 是会对所有的狗产生恐惧还是只对金毛犬产生恐惧?又如何解释上述对“针头”产生恐惧并泛化到“医院”上的现象?因此未来的研究应关注高级认知过程与恐惧泛化的交互作用, 以期完善恐惧泛化的理论基础, 并对相关临床疾病的治疗提供帮助. ...
Fear generalization in humans: Systematic review and implications for anxiety disorder research
2
2015
... 恐惧是一种对环境的适应性机制, 当危险发生时, 恐惧使得机体启动防御机制并对危险刺激进行反应(Dymond, Dunsmoor, Vervliet, Roche, & Hermans, 2015; Hoppenbrouwers, Bulten, & Brazil, 2016; LeDoux, 2000).人类的恐惧反应多是后天习得, 例如, 在华生的条件反射实验中, 小阿尔伯特原本并不惧怕白鼠, 然而当白鼠与巨大的金属响声(危险刺激)同时出现后, 小阿尔伯特便对白鼠产生了恐惧反应, 此后当他再见到小白兔、白色绒毛玩具、甚至老人的白胡须等相关事物时, 他都会感到害怕(Watson & Rayner, 1920).这种不仅对特定的刺激(白鼠)产生恐惧, 而且对与特定刺激相关的事物同样会产生恐惧的现象即是“恐惧泛化” (Dunsmoor, Prince, Murty, Kragel, & LaBar, 2011; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014; Onat & Büchel, 2015).一般来说, 适当的恐惧泛化对个体生存具有积极意义, 因为在漫长的进化过程中, 恐惧泛化能够促使人类躲避各种潜在威胁, 有利于个体生存繁衍; 而在日常生活中, 恐惧泛化也可增强个体的警觉性, 从而有效并快速地对危险环境做出预测和应对(Steimer, 2002). ...
... 为了避免受到既有经验的影响, 实验室研究通常采用简单的感觉信息(如纯音、简单几何图形、面孔)作为实验材料, 同时结合电击、恐惧的面孔、厌恶的气味等负性刺激进行基于知觉的恐惧泛化测试(Dunsmoor & Paz, 2015; Dymond et al., 2015).根据知觉信息的不同维度, 可分为视觉、听觉、情景三方面, 以下将逐一论述. ...
The spread of fear: Symbolic generalization mediates graded threat-avoidance in specific phobia
1
2014
... 从研究内容上来说, 实验室所采用的刺激都是单一的感觉线索, 如图形大小、颜色、声调等, 而在现实生活中的恐惧习得却包含了大量的复杂刺激, 这些刺激通常是多维的, 比如战场、猛兽等.可以说, 基于知觉的恐惧泛化研究低估了现实生活中恐惧泛化的复杂性.另外, 人们常常会根据经验, 去归纳、推断情绪事件的未知属性以及因果联系(Dunsmoor & Murphy, 2015).例如, 一个人对针头产生了恐惧经验, 他就可能对包括医院、护士等在概念上相关的事物产生害怕心理.因此, 基于知觉的恐惧泛化及其相关理论并不能完全解释人类恐惧泛化的现象.例如, Dunsmoor和Murphy (2014)探索了概念的典型性对恐惧泛化的影响, 结果发现对典型成员习得的恐惧会泛化到非典型成员身上, 但非典型成员习得的恐惧则不能泛化到典型成员身上.虽然已有少量研究注意到了如语义、类别、符号等基于高级认知过程的恐惧泛化现象(Bennett, Hermans, Dymond, Vervoort, & Baeyens, 2015; Dunsmoor, White, & LaBar, 2011; Dymond, Schlund, Roche, & Whelan, 2014), 但这方面的实证研究仍然很薄弱, 尚有许多实际生活中的问题不能得到合理解释, 如一个人被金毛犬咬过之后, 是会对所有的狗产生恐惧还是只对金毛犬产生恐惧?又如何解释上述对“针头”产生恐惧并泛化到“医院”上的现象?因此未来的研究应关注高级认知过程与恐惧泛化的交互作用, 以期完善恐惧泛化的理论基础, 并对相关临床疾病的治疗提供帮助. ...
Oxytocin facilitates the extinction of conditioned fear in humans
1
2015
... 鉴于恐惧泛化几乎是所有焦虑障碍的主要症状, 而女性又是焦虑障碍的易感人群 (McLean, Asnaani, Litz, & Hofmann, 2011), 因此分析女性激素对恐惧泛化的影响具有重要的临床意义.研究表明, 恐惧泛化的性别差异很可能是由雌二醇导致(Lynch et al., 2014), 雌二醇或通过促进海马的“模式完成”或者损害“模式分离”来扩大恐惧泛化(Lynch, Winiecki, Vanderhoof, Riccio, & Jasnow, 2016).此外, 催产素有减缓焦虑情绪的作用, 因此可能对治疗焦虑症有重要价值.近期科学家甚至发现催产素可以降低人类和动物的恐惧反应, 促进恐惧的消退(Eckstein et al., 2015; Hyun, Perry, Ganella, & Madsen, 2017; Missig, Ayers, Schulkin, & Rosen, 2010).如把催产素注射到啮齿动物的中央杏仁核(central amygdala)或者背外侧隔膜(dorso-lateral septum)可以减少在消退阶段的恐惧反应(Viviani & Stoop, 2008; Zoicas, Slattery, & Neumann, 2014).在对人类进行的研究中, 使用鼻喷催产素后, 被试mPFC的活动明显增强从而可以达到促使恐惧消退的效果. ...
Noradrenergic blockade stabilizes prefrontal activity and enables fear extinction under stress
1
2015
... 此外, 去甲肾上腺素(norepinephrine, NE)也已成功应用于PTSD的治疗与防治(Brunet et al., 2008; Pervanidou & Chrousos, 2010).动物实验表明, 去甲肾上腺素可能通过调节小鼠内侧前额叶皮层(mPFC)参与调节恐惧的回路, 从而对消退恐惧障碍起到改善作用(Fitzgerald, Giustino, Seemann, & Maren, 2015). ...
A century of generalization
1
2003
... 大量恐惧泛化的研究表明, 相同知觉维度的刺激可以根据物理上的相似性由高到低被分成几个不同的层次, 而个体的恐惧反应也随着刺激的相似性高低而呈现出不同反应大小的梯度 (Ghirlanda & Enquist, 2003; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014).在知觉恐惧泛化研究中, 由一个感觉刺激(记为CS+; 如特定的灯光、声音、形状)与恐惧刺激(US; 如电击、恐惧图片、尖叫声)建立的条件反应, 另一个区别于CS+的感觉刺激作为对照刺激从不匹配US (记为CS-; 如与CS+差别较大的灯光、声音、形状), 通常用相同知觉维度的刺激(记为GS; 如在CS+和CS-之间变化的灯光、声音、形状)进行泛化测试(Hermans, Baeyens, & Vervliet., 2013).这种基于知觉的恐惧泛化梯度可以反映辨别学习和恐惧的泛化程度(Lissek et al., 2008; Struyf, Zaman, Vervliet, & van Diest, 2015) (见图1). ...
Neuronal encoding of the switch from specific to generalized fear
1
2015
... 听觉是仅次于视觉的重要感觉通道, 也是动物和人类感知环境、逃避危险的线索之一.在恐惧泛化的研究中, 研究者们根据频率把声音划分成等距且可辨别的几段音频, 并与电击匹配, 来研究听觉恐惧泛化的机制.动物研究中对恐惧泛化脑机制的探索通常采用声音辨别任务(Ghosh & Chattarji, 2015; Resnik & Paz, 2015).Hovland等(1937)首次将该方法用以研究人类的恐惧泛化, 实验采用4个振幅相同、声音频率等距且可辨别的纯音作为材料, 结果发现被试对从未与US匹配的其他三个纯音也会产生恐惧反应.此外, 研究者用声音结合不同强度的电击, 探讨不同强度电击对恐惧泛化的影响, 结果表明, 不同强度的电击对被试习得恐惧并没有产生影响, 但高强度的电击却可以增加泛化的范围(Dunsmoor, Kroes, Braren, & Phelps, 2017). ...
Dissociating hippocampal subregions: A double dissociation between dentate gyrus and CA1
1
2001
... 海马中 “模式分离(pattern separation)”和“模式完成(pattern completion)”负责对刺激的属性特征与已存储的信息进行匹配(Gluck & Myers, 1993).“模式分离”由海马的齿状回执行(dentate gyrus, DG), 是一种把相似信息区分开的神经活动; 而“模式完成”主要发生在海马CA3区, 它描述了相反的加工过程, 如果刺激的区别较小, “模式完成”则会忽略刺激间的区别(Drew & Huckleberry, 2017; Gilbert, Kesner, & Lee, 2001; Treves & Rolls, 1992).在知觉泛化过程中, 当GS和CS相似性较高时, 海马启动“模式完成”使CS的神经表征激活, 从而进一步引发被试的恐惧反应; 而当GS和CS相似性较低时, 海马则启动“模式分离”激活腹内测前额叶从而降低杏仁核的活动, 达到抑制恐惧反应的效果(Lissek, 2012; Lissek, Bradford et al., 2014).例如, 研究发现在啮齿动物的情景辨别恐惧泛化任务中, 海马CA3区的神经抑制会促进啮齿动物在恐惧和中性情景下的“模式完成”过程, 从而增加恐惧的泛化; 反过来, 齿状回处的神经兴奋则会促进“模式分离”过程, 以抑制恐惧泛化(Sahay et al., 2011; Niibori et al., 2012).可见, 海马在知觉恐惧泛化的过程中负责刺激的辨别加工. ...
The development of fear learning and generalization in 8-13 year-olds
1
2012
... 第三类研究则用两张中性的面孔作为CS+和CS-, 将恐惧表情的面孔与尖叫声同时作为US, 来探索儿童和青少年群体的恐惧泛化规律.Lau等(2008)最先采用面孔范式对12~13岁患焦虑症的青少年进行恐惧泛化研究, 结果发现患焦虑症的青少年的泛化程度较正常组更为严重.随后, Glenn等 (2012)用该范式对8~10岁、11~13岁两个年龄阶段的儿童进行恐惧泛化的测试, 结果发现, 年长被试能够较准确地习得恐惧且泛化梯度较有规律; 而年幼被试则在泛化阶段表现出对CS-而非GS更强的恐惧反应, 这说明年长儿童对危险刺激更具有分辨力.此外, Schiele等(2016)通过对比儿童和成人的恐惧泛化, 发现儿童较成人在唤醒度和皮肤电上都显示出了更多的泛化.Schieler认为泛化梯度具有由浅入深的发展性, 很可能与大脑结构的成熟度有关, 并且大脑结构的成熟度可以调节对危险和安全线索的辨别. ...
Hippocampal mediation of stimulus representation: A computational theory
1
1993
... 海马中 “模式分离(pattern separation)”和“模式完成(pattern completion)”负责对刺激的属性特征与已存储的信息进行匹配(Gluck & Myers, 1993).“模式分离”由海马的齿状回执行(dentate gyrus, DG), 是一种把相似信息区分开的神经活动; 而“模式完成”主要发生在海马CA3区, 它描述了相反的加工过程, 如果刺激的区别较小, “模式完成”则会忽略刺激间的区别(Drew & Huckleberry, 2017; Gilbert, Kesner, & Lee, 2001; Treves & Rolls, 1992).在知觉泛化过程中, 当GS和CS相似性较高时, 海马启动“模式完成”使CS的神经表征激活, 从而进一步引发被试的恐惧反应; 而当GS和CS相似性较低时, 海马则启动“模式分离”激活腹内测前额叶从而降低杏仁核的活动, 达到抑制恐惧反应的效果(Lissek, 2012; Lissek, Bradford et al., 2014).例如, 研究发现在啮齿动物的情景辨别恐惧泛化任务中, 海马CA3区的神经抑制会促进啮齿动物在恐惧和中性情景下的“模式完成”过程, 从而增加恐惧的泛化; 反过来, 齿状回处的神经兴奋则会促进“模式分离”过程, 以抑制恐惧泛化(Sahay et al., 2011; Niibori et al., 2012).可见, 海马在知觉恐惧泛化的过程中负责刺激的辨别加工. ...
a). Neural reactivity tracks fear generalization gradients
2
2013
... 脑岛和海马、杏仁核、水管周灰质(periaqueductal gray )共同控制自主神经系统, 它们在加工恐惧情绪时被激活(Saper, 1982).前人研究表明, 当个体面对不确定的情景及对恐惧刺激进行预期时, 脑岛的响应随之增强(Berns et al., 2006; Dunsmoor, Bandettini, & Knight, 2007).一般认为, 前脑岛是躯体感觉和内脏感觉的结合点, 与ACC、杏仁核、辅助运动区(supplementary motor area, SMA)等区域一起引发与恐惧相关的行为.Dunsmoor, Prince等(2011)首次用功能成像技术结合情绪面孔感知维度对人类恐惧泛化进行研究, 并证明了在条件性恐惧的习得和表达中, 当GS接近CS时, 脑岛、尾状核(caudate)、丘脑的激活程度普遍增加.例如Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013a)在用几何图形测试人类恐惧泛化的过程中, 发现在接近CS时, 脑岛、尾状核、ACC、SMA的响应会随着刺激相似性的增加而增强.随后, Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013b)用相同的范式对广泛性焦虑障碍患者进行恐惧泛化测试, 发现前脑岛(anterior insular, AI)与ACC、杏仁核、SMA连接增强.同时, Lissek, Bradford 等(2014)用圆圈刺激发现在恐惧泛化任务中, 正常被试脑岛激活增强.此后, Onat和Büchel (2015)用面孔范式发现脑岛对CS+最敏感, 颞下皮层(inferotemporal cortex, ITC)对CS+和CS-中间的模糊刺激比较敏感, 这表明前脑岛皮层编码“危险识别”过程, 颞下皮层则对“模糊不清”的信息进行编码.可见, 脑岛在知觉的恐惧泛化过程中参与恐惧的表达和刺激的辨别. ...
... 内侧前额叶皮层(medial prefrontal cortex, mPFC)是恐惧神经环路的关键脑区, 在恐惧消退过程中起着抑制恐惧的作用(Milad et al., 2007; Orsini & Maren, 2012).近来研究表明, mPFC与恐惧泛化过程也有密切的联系.Cullen, Gilman, Winiecki, Riccio和Jasnow (2015)发现, 情景泛化记忆的表达与前扣带皮层(anterior cingulate cortex, ACC)、下边缘皮层(infralimbic cortex, IL)、前边缘皮层(prelimbic cortex, PL)这些前额叶皮层脑区的活性有关, 其中ACC与海马CA1区的连接抑制将减弱恐惧的泛化.另外, 与脑岛等脑区相反的是, vmPFC在接近CS-时激活更强(Greenberg et al., 2013a, 2013b; Lissek, Brandford et al., 2014), 在安全线索条件下GAD患者的vmPFC激活较正常人弱(Greenberg et al., 2013b).此外, vmPFC皮质厚度和恐惧泛化梯度有关, vmPFC皮质的减少可能是导致GAD患者过度泛化的原因之一(Cha et al, 2014).Xu等(2012)观察到mPFC处突触传递的毁损将导致动物情景恐惧记忆的过度泛化.他们认为mPFC、海马以及连接核(nucleus reuniens, NR) (丘脑中线核群的一部分)共同控制着恐惧记忆的泛化.具体而言, 由于mPFC是参与动机及情绪状态的关键脑区, 因此mPFC-NR路径将记忆的动机以及情绪效价信息传递至海马进行特征的重叠编码, 海马又将编码后的信息返回至mPFC进而对泛化记忆进行调节(Xu & Südhof, 2013).另有研究表明, 老鼠对声音习得的恐惧反应和mPFC处胆碱能系统(nitrergic system) (神经元轴突分泌的一类神经递质)的激活水平能够预测泛化的程度, 表现为胆碱能系统激活程度越高, 泛化程度越高(Saul’skaya & Sudorgina, 2016).此外, Likhtik, Stujenske, Topiwala, Harris和Gordon (2014)认为内侧前额叶(mPFC)以及基底外侧核(BLA)连接的增强预示着内侧前额叶可能会影响基底外侧核的活动, 并通过抑制杏仁核的活动来减弱恐惧反应.相反的, mPFC-BLA连接的减弱则会导致对杏仁核的抑制失败, 从而增强恐惧反应.可见, mPFC在知觉恐惧泛化过程中参与调节和抑制恐惧. ...
b). Ventromedial prefrontal cortex reactivity is altered in generalized anxiety disorder during fear generalization
3
2013
... 脑岛和海马、杏仁核、水管周灰质(periaqueductal gray )共同控制自主神经系统, 它们在加工恐惧情绪时被激活(Saper, 1982).前人研究表明, 当个体面对不确定的情景及对恐惧刺激进行预期时, 脑岛的响应随之增强(Berns et al., 2006; Dunsmoor, Bandettini, & Knight, 2007).一般认为, 前脑岛是躯体感觉和内脏感觉的结合点, 与ACC、杏仁核、辅助运动区(supplementary motor area, SMA)等区域一起引发与恐惧相关的行为.Dunsmoor, Prince等(2011)首次用功能成像技术结合情绪面孔感知维度对人类恐惧泛化进行研究, 并证明了在条件性恐惧的习得和表达中, 当GS接近CS时, 脑岛、尾状核(caudate)、丘脑的激活程度普遍增加.例如Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013a)在用几何图形测试人类恐惧泛化的过程中, 发现在接近CS时, 脑岛、尾状核、ACC、SMA的响应会随着刺激相似性的增加而增强.随后, Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013b)用相同的范式对广泛性焦虑障碍患者进行恐惧泛化测试, 发现前脑岛(anterior insular, AI)与ACC、杏仁核、SMA连接增强.同时, Lissek, Bradford 等(2014)用圆圈刺激发现在恐惧泛化任务中, 正常被试脑岛激活增强.此后, Onat和Büchel (2015)用面孔范式发现脑岛对CS+最敏感, 颞下皮层(inferotemporal cortex, ITC)对CS+和CS-中间的模糊刺激比较敏感, 这表明前脑岛皮层编码“危险识别”过程, 颞下皮层则对“模糊不清”的信息进行编码.可见, 脑岛在知觉的恐惧泛化过程中参与恐惧的表达和刺激的辨别. ...
... 内侧前额叶皮层(medial prefrontal cortex, mPFC)是恐惧神经环路的关键脑区, 在恐惧消退过程中起着抑制恐惧的作用(Milad et al., 2007; Orsini & Maren, 2012).近来研究表明, mPFC与恐惧泛化过程也有密切的联系.Cullen, Gilman, Winiecki, Riccio和Jasnow (2015)发现, 情景泛化记忆的表达与前扣带皮层(anterior cingulate cortex, ACC)、下边缘皮层(infralimbic cortex, IL)、前边缘皮层(prelimbic cortex, PL)这些前额叶皮层脑区的活性有关, 其中ACC与海马CA1区的连接抑制将减弱恐惧的泛化.另外, 与脑岛等脑区相反的是, vmPFC在接近CS-时激活更强(Greenberg et al., 2013a, 2013b; Lissek, Brandford et al., 2014), 在安全线索条件下GAD患者的vmPFC激活较正常人弱(Greenberg et al., 2013b).此外, vmPFC皮质厚度和恐惧泛化梯度有关, vmPFC皮质的减少可能是导致GAD患者过度泛化的原因之一(Cha et al, 2014).Xu等(2012)观察到mPFC处突触传递的毁损将导致动物情景恐惧记忆的过度泛化.他们认为mPFC、海马以及连接核(nucleus reuniens, NR) (丘脑中线核群的一部分)共同控制着恐惧记忆的泛化.具体而言, 由于mPFC是参与动机及情绪状态的关键脑区, 因此mPFC-NR路径将记忆的动机以及情绪效价信息传递至海马进行特征的重叠编码, 海马又将编码后的信息返回至mPFC进而对泛化记忆进行调节(Xu & Südhof, 2013).另有研究表明, 老鼠对声音习得的恐惧反应和mPFC处胆碱能系统(nitrergic system) (神经元轴突分泌的一类神经递质)的激活水平能够预测泛化的程度, 表现为胆碱能系统激活程度越高, 泛化程度越高(Saul’skaya & Sudorgina, 2016).此外, Likhtik, Stujenske, Topiwala, Harris和Gordon (2014)认为内侧前额叶(mPFC)以及基底外侧核(BLA)连接的增强预示着内侧前额叶可能会影响基底外侧核的活动, 并通过抑制杏仁核的活动来减弱恐惧反应.相反的, mPFC-BLA连接的减弱则会导致对杏仁核的抑制失败, 从而增强恐惧反应.可见, mPFC在知觉恐惧泛化过程中参与调节和抑制恐惧. ...
... ), 在安全线索条件下GAD患者的vmPFC激活较正常人弱(Greenberg et al., 2013b).此外, vmPFC皮质厚度和恐惧泛化梯度有关, vmPFC皮质的减少可能是导致GAD患者过度泛化的原因之一(Cha et al, 2014).Xu等(2012)观察到mPFC处突触传递的毁损将导致动物情景恐惧记忆的过度泛化.他们认为mPFC、海马以及连接核(nucleus reuniens, NR) (丘脑中线核群的一部分)共同控制着恐惧记忆的泛化.具体而言, 由于mPFC是参与动机及情绪状态的关键脑区, 因此mPFC-NR路径将记忆的动机以及情绪效价信息传递至海马进行特征的重叠编码, 海马又将编码后的信息返回至mPFC进而对泛化记忆进行调节(Xu & Südhof, 2013).另有研究表明, 老鼠对声音习得的恐惧反应和mPFC处胆碱能系统(nitrergic system) (神经元轴突分泌的一类神经递质)的激活水平能够预测泛化的程度, 表现为胆碱能系统激活程度越高, 泛化程度越高(Saul’skaya & Sudorgina, 2016).此外, Likhtik, Stujenske, Topiwala, Harris和Gordon (2014)认为内侧前额叶(mPFC)以及基底外侧核(BLA)连接的增强预示着内侧前额叶可能会影响基底外侧核的活动, 并通过抑制杏仁核的活动来减弱恐惧反应.相反的, mPFC-BLA连接的减弱则会导致对杏仁核的抑制失败, 从而增强恐惧反应.可见, mPFC在知觉恐惧泛化过程中参与调节和抑制恐惧. ...
Generalization of acquired emotional responses
1
2013
... 大量恐惧泛化的研究表明, 相同知觉维度的刺激可以根据物理上的相似性由高到低被分成几个不同的层次, 而个体的恐惧反应也随着刺激的相似性高低而呈现出不同反应大小的梯度 (Ghirlanda & Enquist, 2003; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014).在知觉恐惧泛化研究中, 由一个感觉刺激(记为CS+; 如特定的灯光、声音、形状)与恐惧刺激(US; 如电击、恐惧图片、尖叫声)建立的条件反应, 另一个区别于CS+的感觉刺激作为对照刺激从不匹配US (记为CS-; 如与CS+差别较大的灯光、声音、形状), 通常用相同知觉维度的刺激(记为GS; 如在CS+和CS-之间变化的灯光、声音、形状)进行泛化测试(Hermans, Baeyens, & Vervliet., 2013).这种基于知觉的恐惧泛化梯度可以反映辨别学习和恐惧的泛化程度(Lissek et al., 2008; Struyf, Zaman, Vervliet, & van Diest, 2015) (见图1). ...
A parametric study of fear generalization to faces and non-face objects: Relationship to discrimination thresholds
2
2014
... 第一类研究通常以两张同性别个体的中性面孔作为CS+和CS-, GS则是在CS+和CS-之间融合的面孔.如, Onat和Büchel (2015)运用该范式发现被试的皮肤电水平随着面孔的相似性增加而增强.另外, Holt等(2014)采用该法并结合差别域限法(JND)设置GS.GS被分成能够察觉的面孔和不能察觉的面孔两大类.结果表明, 阈限下的面孔比阈限上的面孔能引发更大的恐惧反应. ...
... 从研究方法上来说, 刺激的不同设置方式是否会影响被试的辨别力?首先, 在对GS恐惧程度进行评估时, 刺激单独呈现, 并被较长的时间间隔分开, 这就减弱了刺激间直接的对比.此外, 不同的时间间隔将会触发不同的记忆过程, 因为随着时间间隔的增长, 被试的行为反应会相应减弱(Burton & Sinclair, 2000).其次, 被试的情绪状态也会影响实验结果.在单纯的知觉辨别任务中, 没有呈现厌恶刺激时被试的情绪是相对中性的.但在恐惧泛化任务中, 恐惧的情绪状态会影响被试的决策并减少知觉的准确性, 从而影响被试对GS和CS的辨别(Stefanucci & Storbeck, 2009).由此可见, 个体在恐惧状态和正常状态下的辨别能力是存在差异的.再次, 刺激梯度通常是人为操纵, 而非基于被试的知觉, 而个体的感知阈限不同, 对刺激的辨别度也存在差异, 如Resnik, Sobel和Paz (2011)用纯音(1/2 kHz)分别作为CS+和CS-, 被试却经常把CS+周围的刺激认作CS+.然而, 只有极少数研究用差别阈限法关注被试对材料的辨别度(Holt et al., 2014).所以未来的研究应该注意区分被试对刺激的辨别力, 采用差别阈限法对实验材料进行设置以避免因混淆辨别力不佳而造成的实验结果误差. ...
Parsing fear: A reassessment of the evidence for fear deficits in psychopathy
1
2016
... 恐惧是一种对环境的适应性机制, 当危险发生时, 恐惧使得机体启动防御机制并对危险刺激进行反应(Dymond, Dunsmoor, Vervliet, Roche, & Hermans, 2015; Hoppenbrouwers, Bulten, & Brazil, 2016; LeDoux, 2000).人类的恐惧反应多是后天习得, 例如, 在华生的条件反射实验中, 小阿尔伯特原本并不惧怕白鼠, 然而当白鼠与巨大的金属响声(危险刺激)同时出现后, 小阿尔伯特便对白鼠产生了恐惧反应, 此后当他再见到小白兔、白色绒毛玩具、甚至老人的白胡须等相关事物时, 他都会感到害怕(Watson & Rayner, 1920).这种不仅对特定的刺激(白鼠)产生恐惧, 而且对与特定刺激相关的事物同样会产生恐惧的现象即是“恐惧泛化” (Dunsmoor, Prince, Murty, Kragel, & LaBar, 2011; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014; Onat & Büchel, 2015).一般来说, 适当的恐惧泛化对个体生存具有积极意义, 因为在漫长的进化过程中, 恐惧泛化能够促使人类躲避各种潜在威胁, 有利于个体生存繁衍; 而在日常生活中, 恐惧泛化也可增强个体的警觉性, 从而有效并快速地对危险环境做出预测和应对(Steimer, 2002). ...
The generalization of conditioned responses: I. The sensory generalization of conditioned responses with varying frequencies of tone
1
1937
... 听觉是仅次于视觉的重要感觉通道, 也是动物和人类感知环境、逃避危险的线索之一.在恐惧泛化的研究中, 研究者们根据频率把声音划分成等距且可辨别的几段音频, 并与电击匹配, 来研究听觉恐惧泛化的机制.动物研究中对恐惧泛化脑机制的探索通常采用声音辨别任务(Ghosh & Chattarji, 2015; Resnik & Paz, 2015).Hovland等(1937)首次将该方法用以研究人类的恐惧泛化, 实验采用4个振幅相同、声音频率等距且可辨别的纯音作为材料, 结果发现被试对从未与US匹配的其他三个纯音也会产生恐惧反应.此外, 研究者用声音结合不同强度的电击, 探讨不同强度电击对恐惧泛化的影响, 结果表明, 不同强度的电击对被试习得恐惧并没有产生影响, 但高强度的电击却可以增加泛化的范围(Dunsmoor, Kroes, Braren, & Phelps, 2017). ...
Postnatal development of neurotransmitter systems and their relevance to extinction of conditioned fear
1
2017
... 鉴于恐惧泛化几乎是所有焦虑障碍的主要症状, 而女性又是焦虑障碍的易感人群 (McLean, Asnaani, Litz, & Hofmann, 2011), 因此分析女性激素对恐惧泛化的影响具有重要的临床意义.研究表明, 恐惧泛化的性别差异很可能是由雌二醇导致(Lynch et al., 2014), 雌二醇或通过促进海马的“模式完成”或者损害“模式分离”来扩大恐惧泛化(Lynch, Winiecki, Vanderhoof, Riccio, & Jasnow, 2016).此外, 催产素有减缓焦虑情绪的作用, 因此可能对治疗焦虑症有重要价值.近期科学家甚至发现催产素可以降低人类和动物的恐惧反应, 促进恐惧的消退(Eckstein et al., 2015; Hyun, Perry, Ganella, & Madsen, 2017; Missig, Ayers, Schulkin, & Rosen, 2010).如把催产素注射到啮齿动物的中央杏仁核(central amygdala)或者背外侧隔膜(dorso-lateral septum)可以减少在消退阶段的恐惧反应(Viviani & Stoop, 2008; Zoicas, Slattery, & Neumann, 2014).在对人类进行的研究中, 使用鼻喷催产素后, 被试mPFC的活动明显增强从而可以达到促使恐惧消退的效果. ...
Perspectives on fear generalization and its implications for emotional disorders
2
2017
... 情景的恐惧泛化需要对整个情景内的信息进行辨别, 这些信息通常包括空间、嗅觉、听觉和视觉信息, 而不是单一的物体特征的变化(Jasnow, Lynch, Gilman, & Riccio, 2017).首先, 在动物研究方面, 有研究者训练啮齿类动物对某一情景(CXT+)习得恐惧, 而在另外一个新异的情景(CXT-)下始终不匹配恐惧刺激, 紧接着在一个和CXT+相似的情景(G-CXT)中进行恐惧泛化测试.这些情景任务包括辨别触摸屏中物体与屏幕的距离(Mcallister, Saksida, & Bussey, 2013; Talpos, McTighe, Dias, Saksida, & Bussey, 2010)、辨别新异刺激和熟悉刺激的位置(Bekinschtein et al., 2013)、在不同形状的空间里习得恐惧(Mchugh et al., 2007)等.动物研究表明, 恐惧习得和重现刺激情景(G-CXT)之间的时间间隔越长, 恐惧泛化程度越大(Biedenkapp & Rudy, 2007).而在习得任务之前预先暴露情景则会减弱恐惧的泛化.此外, 研究者通过运用情景恐惧泛化范式, 发现啮齿动物恐惧泛化存在着性别差异, 具体表现为雌性比雄性更容易发生泛化(Keiser et al., 2017), 且泛化速度更快(Lynch, Cullen, Jasnow, & Riccio, 2013).预先暴露情景增加了雄性老鼠在习得阶段的恐惧反应, 但却减少了雌性老鼠在泛化的阶段恐惧反应(Keiser et al., 2017). ...
... 杏仁核是恐惧学习和恐惧泛化的关键脑区, 是CS与US的汇聚点并启动由威胁引起的条件反应.运用情景辨别任务, Ciocchi等(2010)发现激活中央杏仁核外侧核(lateral part of central amygdala, CeL)神经元的活动, 或者抑制中央杏仁核内侧核(medial part of central amygdala, CeM)神经元的活动都会增加老鼠对CS-的恐惧反应.这说明杏仁核中央核团(central amygdala, CeA)的神经活性对恐惧泛化具有一定的调节作用.Lopresto, Schipper和Homberg (2016)认为探索杏仁核在恐惧泛化中的作用即是把杏仁核看作神经回路的节点, 该节点连接着海马和大脑皮层.在情景任务中, 已有研究表明恐惧情景引发老鼠和人类的杏仁核以及海马的响应比中性情景更强(Maren, Phan, & Liberzon, 2013; Marschner, Kalisch, Vervliet, Vansteenwegen, & Buchel, 2008).这说明, 杏仁核以及海马可能是调节情景恐惧泛化的关键脑区之一.此后, Resnik和Paz (2015)用纯音作为刺激材料考察了猴子恐惧泛化的神经机制及其行为反应, 发现基底外侧核(basolateral complex of the amygdala, BLA)处神经元的特异变化减弱了泛化阶段的辨别学习, 这种杏仁核部位的轻微改变影响了前额叶皮层以及海马, 并破坏了泛化的平衡, 从而导致了更广的泛化.另外, 内侧前额叶(mPFC)-BLA通路的增强可以促进恐惧消退, 这种自上而下连接不良的传递, 也会导致更广的泛化(Jasnow et al., 2017; Likhtik & Paz, 2015).由此可见, 杏仁核在知觉恐惧泛化回路中有着重要的作用, 是其他脑区与感知觉信息的连接点, 控制着恐惧反应的表达. ...
Sex differences in context fear generalization and recruitment of hippocampus and amygdala during retrieval
2
2017
... 情景的恐惧泛化需要对整个情景内的信息进行辨别, 这些信息通常包括空间、嗅觉、听觉和视觉信息, 而不是单一的物体特征的变化(Jasnow, Lynch, Gilman, & Riccio, 2017).首先, 在动物研究方面, 有研究者训练啮齿类动物对某一情景(CXT+)习得恐惧, 而在另外一个新异的情景(CXT-)下始终不匹配恐惧刺激, 紧接着在一个和CXT+相似的情景(G-CXT)中进行恐惧泛化测试.这些情景任务包括辨别触摸屏中物体与屏幕的距离(Mcallister, Saksida, & Bussey, 2013; Talpos, McTighe, Dias, Saksida, & Bussey, 2010)、辨别新异刺激和熟悉刺激的位置(Bekinschtein et al., 2013)、在不同形状的空间里习得恐惧(Mchugh et al., 2007)等.动物研究表明, 恐惧习得和重现刺激情景(G-CXT)之间的时间间隔越长, 恐惧泛化程度越大(Biedenkapp & Rudy, 2007).而在习得任务之前预先暴露情景则会减弱恐惧的泛化.此外, 研究者通过运用情景恐惧泛化范式, 发现啮齿动物恐惧泛化存在着性别差异, 具体表现为雌性比雄性更容易发生泛化(Keiser et al., 2017), 且泛化速度更快(Lynch, Cullen, Jasnow, & Riccio, 2013).预先暴露情景增加了雄性老鼠在习得阶段的恐惧反应, 但却减少了雌性老鼠在泛化的阶段恐惧反应(Keiser et al., 2017). ...
... ).预先暴露情景增加了雄性老鼠在习得阶段的恐惧反应, 但却减少了雌性老鼠在泛化的阶段恐惧反应(Keiser et al., 2017). ...
Neural correlates of disgust- and fear-conditioned responses
1
2012
... 在实验材料方面, 以往实验通常采用 “厌恶刺激”作为US和CS来进行匹配, 如“高度恼人但不疼”的电击、恐惧的面孔、尖叫声等.但“厌恶刺激”所能引发的仅仅是单纯的恐惧情绪吗?事实上, 厌恶和恐惧本是两种不同的情绪, 二者在面部表情、行为反应、生理反应以及主观体验上均存在差异(Klucken et al., 2012; Woody & Teachman, 2000).因此, 我们可以质疑以往的的研究是否仅能引发的是单纯的恐惧情绪.总的来说, 未来的研究应尽量设法采用仅能真正激发单纯恐惧情绪的刺激.既关注恐惧刺激材料的准确性, 也可尝试多样化的恐惧刺激材料(如恐惧的图片、场景、声音、激光等). ...
Fear and anxiety: Animal models and human cognitive psychophysiology
1
2000
... 巴普洛夫经典条件反射理论认为, 当安全的条件刺激(conditioned stimulus, CS)和一个令人恐惧的无条件刺激(unconditioned stimulus, US)多次匹配(CS-US)以后, 单独呈现条件刺激即能引起人出汗、呼吸加快、心跳加速等条件性恐惧反应(conditioned responses, CR).巴普洛夫经典条件反射已被广泛应用于研究恐惧的习得、表达、消退的研究中(Lang, Davis, & Öhman, 2000; LeDoux, 1996; Milad et al., 2007).然而恐惧学习并不仅限于恐惧条件化情形之下的特定事件或事物, 任何与恐惧刺激相关的事物都有可能引发恐惧反应, 这即是恐惧泛化(fear generalization), 而那些从未与无条件刺激匹配却能够引起恐惧反应的相关刺激便被称为泛化刺激(generalization stimulus, GS). ...
Fear conditioning in adolescents with anxiety disorders: Results from a novel experimental paradigm
1
2008
... 第三类研究则用两张中性的面孔作为CS+和CS-, 将恐惧表情的面孔与尖叫声同时作为US, 来探索儿童和青少年群体的恐惧泛化规律.Lau等(2008)最先采用面孔范式对12~13岁患焦虑症的青少年进行恐惧泛化研究, 结果发现患焦虑症的青少年的泛化程度较正常组更为严重.随后, Glenn等 (2012)用该范式对8~10岁、11~13岁两个年龄阶段的儿童进行恐惧泛化的测试, 结果发现, 年长被试能够较准确地习得恐惧且泛化梯度较有规律; 而年幼被试则在泛化阶段表现出对CS-而非GS更强的恐惧反应, 这说明年长儿童对危险刺激更具有分辨力.此外, Schiele等(2016)通过对比儿童和成人的恐惧泛化, 发现儿童较成人在唤醒度和皮肤电上都显示出了更多的泛化.Schieler认为泛化梯度具有由浅入深的发展性, 很可能与大脑结构的成熟度有关, 并且大脑结构的成熟度可以调节对危险和安全线索的辨别. ...
Behavioral and neural mechanisms of overgeneralization in anxiety
2
2016
... 此外, 也有研究者以声音为材料, 发现情绪效价对恐惧泛化也具有重要影响.例如, Laufer和Paz (2012)对金钱损失和收益条件下被试的泛化程度进行了探讨.结果发现, 与金钱收益条件相比, 金钱损失条件下被试的感受性会明显降低, 知觉阈限升高, 恐惧泛化程度也表现得更广; 这说明, 对金钱损失的恐惧感会降低被试对刺激的辨别能力.近期研究发现, 广泛性焦虑(GAD)患者在赢钱和输钱情况下的泛化情况都比正常人严重(Laufer, Israeli, & Paz, 2016). ...
... Laufer等(2016)用声音刺激发现, 相比于正常被试, GAD被试的dACC、尾状核、初级听觉皮层(primary auditory cortex)激活更强; 而正常被试的vmPFC和腹侧前扣带回 (ventral ACC) 则强于GAD被试.另外GAD被试在习得阶段dACC响应与负性刺激的泛化相关; 而尾状核(putamen)的响应与正性刺激的泛化相关.这表明在恐惧学习阶段的脑区响应对恐惧泛化阶段有一定预测作用.Dunsmoor, Prince等(2011)用面孔刺激发现泛化阶段杏仁核与负责面孔识别的脑区——梭状回(fusiform gyrus, FFG)的连接增强.由此可见, 杏仁核与初级感觉皮层的连接通过提高恐惧相关联的感觉信息来促进恐惧的表达. ...
Monetary loss alters perceptual thresholds and compromises future decisions via amygdala and prefrontal networks
1
2012
... 此外, 也有研究者以声音为材料, 发现情绪效价对恐惧泛化也具有重要影响.例如, Laufer和Paz (2012)对金钱损失和收益条件下被试的泛化程度进行了探讨.结果发现, 与金钱收益条件相比, 金钱损失条件下被试的感受性会明显降低, 知觉阈限升高, 恐惧泛化程度也表现得更广; 这说明, 对金钱损失的恐惧感会降低被试对刺激的辨别能力.近期研究发现, 广泛性焦虑(GAD)患者在赢钱和输钱情况下的泛化情况都比正常人严重(Laufer, Israeli, & Paz, 2016). ...
1
1996
... 巴普洛夫经典条件反射理论认为, 当安全的条件刺激(conditioned stimulus, CS)和一个令人恐惧的无条件刺激(unconditioned stimulus, US)多次匹配(CS-US)以后, 单独呈现条件刺激即能引起人出汗、呼吸加快、心跳加速等条件性恐惧反应(conditioned responses, CR).巴普洛夫经典条件反射已被广泛应用于研究恐惧的习得、表达、消退的研究中(Lang, Davis, & Öhman, 2000; LeDoux, 1996; Milad et al., 2007).然而恐惧学习并不仅限于恐惧条件化情形之下的特定事件或事物, 任何与恐惧刺激相关的事物都有可能引发恐惧反应, 这即是恐惧泛化(fear generalization), 而那些从未与无条件刺激匹配却能够引起恐惧反应的相关刺激便被称为泛化刺激(generalization stimulus, GS). ...
Emotion circuits in the brain
2
2000
... 恐惧是一种对环境的适应性机制, 当危险发生时, 恐惧使得机体启动防御机制并对危险刺激进行反应(Dymond, Dunsmoor, Vervliet, Roche, & Hermans, 2015; Hoppenbrouwers, Bulten, & Brazil, 2016; LeDoux, 2000).人类的恐惧反应多是后天习得, 例如, 在华生的条件反射实验中, 小阿尔伯特原本并不惧怕白鼠, 然而当白鼠与巨大的金属响声(危险刺激)同时出现后, 小阿尔伯特便对白鼠产生了恐惧反应, 此后当他再见到小白兔、白色绒毛玩具、甚至老人的白胡须等相关事物时, 他都会感到害怕(Watson & Rayner, 1920).这种不仅对特定的刺激(白鼠)产生恐惧, 而且对与特定刺激相关的事物同样会产生恐惧的现象即是“恐惧泛化” (Dunsmoor, Prince, Murty, Kragel, & LaBar, 2011; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014; Onat & Büchel, 2015).一般来说, 适当的恐惧泛化对个体生存具有积极意义, 因为在漫长的进化过程中, 恐惧泛化能够促使人类躲避各种潜在威胁, 有利于个体生存繁衍; 而在日常生活中, 恐惧泛化也可增强个体的警觉性, 从而有效并快速地对危险环境做出预测和应对(Steimer, 2002). ...
... 为能够直观地理解恐惧泛化与相关脑区之间的联系, 本文将结合LeDoux (2000)提出的恐惧学习神经机制以及Lissek (2012)提出的恐惧泛化神经机制, 进一步对恐惧泛化神经回路进行描述(见图2).Ledoux认为从刺激呈现到恐惧反应存在两条通路:皮层上的“高通路” (high road)和皮层下的“低通路” (low road), 杏仁核(amygdala)则是连接两条通路的枢纽.其中, “高通路”是一条慢速而精细的通路, 感觉信息先被投射至丘脑(thalamus), 然后丘脑将信息发送至感觉皮质(sensory cortex)进行更细致的分析, 最后感觉皮质再将结果投射至杏仁核.而“低通路”则是一条快速而粗略的通路, 感觉信息投射至丘脑, 丘脑直接将信号传入杏仁核.感觉信息通过“高通路”和“低通路”投射到杏仁核之后, 杏仁核再与脑岛(insula)、脑干(brainstem)建立连接, 从而引发恐惧反应. ...
Reduced hippocampal volume is associated with overgeneralization of negative context in individuals with PTSD
0
2015
Amygdala-prefrontal interactions in (mal)adaptive learning
1
2015
... 杏仁核是恐惧学习和恐惧泛化的关键脑区, 是CS与US的汇聚点并启动由威胁引起的条件反应.运用情景辨别任务, Ciocchi等(2010)发现激活中央杏仁核外侧核(lateral part of central amygdala, CeL)神经元的活动, 或者抑制中央杏仁核内侧核(medial part of central amygdala, CeM)神经元的活动都会增加老鼠对CS-的恐惧反应.这说明杏仁核中央核团(central amygdala, CeA)的神经活性对恐惧泛化具有一定的调节作用.Lopresto, Schipper和Homberg (2016)认为探索杏仁核在恐惧泛化中的作用即是把杏仁核看作神经回路的节点, 该节点连接着海马和大脑皮层.在情景任务中, 已有研究表明恐惧情景引发老鼠和人类的杏仁核以及海马的响应比中性情景更强(Maren, Phan, & Liberzon, 2013; Marschner, Kalisch, Vervliet, Vansteenwegen, & Buchel, 2008).这说明, 杏仁核以及海马可能是调节情景恐惧泛化的关键脑区之一.此后, Resnik和Paz (2015)用纯音作为刺激材料考察了猴子恐惧泛化的神经机制及其行为反应, 发现基底外侧核(basolateral complex of the amygdala, BLA)处神经元的特异变化减弱了泛化阶段的辨别学习, 这种杏仁核部位的轻微改变影响了前额叶皮层以及海马, 并破坏了泛化的平衡, 从而导致了更广的泛化.另外, 内侧前额叶(mPFC)-BLA通路的增强可以促进恐惧消退, 这种自上而下连接不良的传递, 也会导致更广的泛化(Jasnow et al., 2017; Likhtik & Paz, 2015).由此可见, 杏仁核在知觉恐惧泛化回路中有着重要的作用, 是其他脑区与感知觉信息的连接点, 控制着恐惧反应的表达. ...
Prefrontal entrainment of amygdala activity signals safety in learned fear and innate anxiety
1
2014
... 内侧前额叶皮层(medial prefrontal cortex, mPFC)是恐惧神经环路的关键脑区, 在恐惧消退过程中起着抑制恐惧的作用(Milad et al., 2007; Orsini & Maren, 2012).近来研究表明, mPFC与恐惧泛化过程也有密切的联系.Cullen, Gilman, Winiecki, Riccio和Jasnow (2015)发现, 情景泛化记忆的表达与前扣带皮层(anterior cingulate cortex, ACC)、下边缘皮层(infralimbic cortex, IL)、前边缘皮层(prelimbic cortex, PL)这些前额叶皮层脑区的活性有关, 其中ACC与海马CA1区的连接抑制将减弱恐惧的泛化.另外, 与脑岛等脑区相反的是, vmPFC在接近CS-时激活更强(Greenberg et al., 2013a, 2013b; Lissek, Brandford et al., 2014), 在安全线索条件下GAD患者的vmPFC激活较正常人弱(Greenberg et al., 2013b).此外, vmPFC皮质厚度和恐惧泛化梯度有关, vmPFC皮质的减少可能是导致GAD患者过度泛化的原因之一(Cha et al, 2014).Xu等(2012)观察到mPFC处突触传递的毁损将导致动物情景恐惧记忆的过度泛化.他们认为mPFC、海马以及连接核(nucleus reuniens, NR) (丘脑中线核群的一部分)共同控制着恐惧记忆的泛化.具体而言, 由于mPFC是参与动机及情绪状态的关键脑区, 因此mPFC-NR路径将记忆的动机以及情绪效价信息传递至海马进行特征的重叠编码, 海马又将编码后的信息返回至mPFC进而对泛化记忆进行调节(Xu & Südhof, 2013).另有研究表明, 老鼠对声音习得的恐惧反应和mPFC处胆碱能系统(nitrergic system) (神经元轴突分泌的一类神经递质)的激活水平能够预测泛化的程度, 表现为胆碱能系统激活程度越高, 泛化程度越高(Saul’skaya & Sudorgina, 2016).此外, Likhtik, Stujenske, Topiwala, Harris和Gordon (2014)认为内侧前额叶(mPFC)以及基底外侧核(BLA)连接的增强预示着内侧前额叶可能会影响基底外侧核的活动, 并通过抑制杏仁核的活动来减弱恐惧反应.相反的, mPFC-BLA连接的减弱则会导致对杏仁核的抑制失败, 从而增强恐惧反应.可见, mPFC在知觉恐惧泛化过程中参与调节和抑制恐惧. ...
Toward an account of clinical anxiety predicated on basic, neurally mapped mechanisms of Pavlovian fear-learning: The case for conditioned overgeneralization
2
2012
... 海马中 “模式分离(pattern separation)”和“模式完成(pattern completion)”负责对刺激的属性特征与已存储的信息进行匹配(Gluck & Myers, 1993).“模式分离”由海马的齿状回执行(dentate gyrus, DG), 是一种把相似信息区分开的神经活动; 而“模式完成”主要发生在海马CA3区, 它描述了相反的加工过程, 如果刺激的区别较小, “模式完成”则会忽略刺激间的区别(Drew & Huckleberry, 2017; Gilbert, Kesner, & Lee, 2001; Treves & Rolls, 1992).在知觉泛化过程中, 当GS和CS相似性较高时, 海马启动“模式完成”使CS的神经表征激活, 从而进一步引发被试的恐惧反应; 而当GS和CS相似性较低时, 海马则启动“模式分离”激活腹内测前额叶从而降低杏仁核的活动, 达到抑制恐惧反应的效果(Lissek, 2012; Lissek, Bradford et al., 2014).例如, 研究发现在啮齿动物的情景辨别恐惧泛化任务中, 海马CA3区的神经抑制会促进啮齿动物在恐惧和中性情景下的“模式完成”过程, 从而增加恐惧的泛化; 反过来, 齿状回处的神经兴奋则会促进“模式分离”过程, 以抑制恐惧泛化(Sahay et al., 2011; Niibori et al., 2012).可见, 海马在知觉恐惧泛化的过程中负责刺激的辨别加工. ...
... 为能够直观地理解恐惧泛化与相关脑区之间的联系, 本文将结合LeDoux (2000)提出的恐惧学习神经机制以及Lissek (2012)提出的恐惧泛化神经机制, 进一步对恐惧泛化神经回路进行描述(见图2).Ledoux认为从刺激呈现到恐惧反应存在两条通路:皮层上的“高通路” (high road)和皮层下的“低通路” (low road), 杏仁核(amygdala)则是连接两条通路的枢纽.其中, “高通路”是一条慢速而精细的通路, 感觉信息先被投射至丘脑(thalamus), 然后丘脑将信息发送至感觉皮质(sensory cortex)进行更细致的分析, 最后感觉皮质再将结果投射至杏仁核.而“低通路”则是一条快速而粗略的通路, 感觉信息投射至丘脑, 丘脑直接将信号传入杏仁核.感觉信息通过“高通路”和“低通路”投射到杏仁核之后, 杏仁核再与脑岛(insula)、脑干(brainstem)建立连接, 从而引发恐惧反应. ...
Generalization of conditioned fear-potentiated startle in humans: Experimental validation and clinical relevance
3
2008
... 恐惧是一种对环境的适应性机制, 当危险发生时, 恐惧使得机体启动防御机制并对危险刺激进行反应(Dymond, Dunsmoor, Vervliet, Roche, & Hermans, 2015; Hoppenbrouwers, Bulten, & Brazil, 2016; LeDoux, 2000).人类的恐惧反应多是后天习得, 例如, 在华生的条件反射实验中, 小阿尔伯特原本并不惧怕白鼠, 然而当白鼠与巨大的金属响声(危险刺激)同时出现后, 小阿尔伯特便对白鼠产生了恐惧反应, 此后当他再见到小白兔、白色绒毛玩具、甚至老人的白胡须等相关事物时, 他都会感到害怕(Watson & Rayner, 1920).这种不仅对特定的刺激(白鼠)产生恐惧, 而且对与特定刺激相关的事物同样会产生恐惧的现象即是“恐惧泛化” (Dunsmoor, Prince, Murty, Kragel, & LaBar, 2011; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014; Onat & Büchel, 2015).一般来说, 适当的恐惧泛化对个体生存具有积极意义, 因为在漫长的进化过程中, 恐惧泛化能够促使人类躲避各种潜在威胁, 有利于个体生存繁衍; 而在日常生活中, 恐惧泛化也可增强个体的警觉性, 从而有效并快速地对危险环境做出预测和应对(Steimer, 2002). ...
... 大量恐惧泛化的研究表明, 相同知觉维度的刺激可以根据物理上的相似性由高到低被分成几个不同的层次, 而个体的恐惧反应也随着刺激的相似性高低而呈现出不同反应大小的梯度 (Ghirlanda & Enquist, 2003; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014).在知觉恐惧泛化研究中, 由一个感觉刺激(记为CS+; 如特定的灯光、声音、形状)与恐惧刺激(US; 如电击、恐惧图片、尖叫声)建立的条件反应, 另一个区别于CS+的感觉刺激作为对照刺激从不匹配US (记为CS-; 如与CS+差别较大的灯光、声音、形状), 通常用相同知觉维度的刺激(记为GS; 如在CS+和CS-之间变化的灯光、声音、形状)进行泛化测试(Hermans, Baeyens, & Vervliet., 2013).这种基于知觉的恐惧泛化梯度可以反映辨别学习和恐惧的泛化程度(Lissek et al., 2008; Struyf, Zaman, Vervliet, & van Diest, 2015) (见图1). ...
... ).这种基于知觉的恐惧泛化梯度可以反映辨别学习和恐惧的泛化程度(Lissek et al., 2008; Struyf, Zaman, Vervliet, & van Diest, 2015) (见图1). ...
Neural substrates of classically conditioned fear-generalization in humans: A parametric fMRI Study
4
2014
... 海马中 “模式分离(pattern separation)”和“模式完成(pattern completion)”负责对刺激的属性特征与已存储的信息进行匹配(Gluck & Myers, 1993).“模式分离”由海马的齿状回执行(dentate gyrus, DG), 是一种把相似信息区分开的神经活动; 而“模式完成”主要发生在海马CA3区, 它描述了相反的加工过程, 如果刺激的区别较小, “模式完成”则会忽略刺激间的区别(Drew & Huckleberry, 2017; Gilbert, Kesner, & Lee, 2001; Treves & Rolls, 1992).在知觉泛化过程中, 当GS和CS相似性较高时, 海马启动“模式完成”使CS的神经表征激活, 从而进一步引发被试的恐惧反应; 而当GS和CS相似性较低时, 海马则启动“模式分离”激活腹内测前额叶从而降低杏仁核的活动, 达到抑制恐惧反应的效果(Lissek, 2012; Lissek, Bradford et al., 2014).例如, 研究发现在啮齿动物的情景辨别恐惧泛化任务中, 海马CA3区的神经抑制会促进啮齿动物在恐惧和中性情景下的“模式完成”过程, 从而增加恐惧的泛化; 反过来, 齿状回处的神经兴奋则会促进“模式分离”过程, 以抑制恐惧泛化(Sahay et al., 2011; Niibori et al., 2012).可见, 海马在知觉恐惧泛化的过程中负责刺激的辨别加工. ...
... Lissek, Bradford等(2014)用大小圆圈作为刺激, 发现人类腹侧海马(ventral hippocampus, VH)的响应会随着刺激间相似性的减弱而增强, 这和“模式分离”加工一致.此外, 海马的体积也会影响恐惧泛化.如Levy-Gigi, Szabo, Richter-Levin和Kéri (2014)发现PTSD患者海马体积的减少和负性情景的泛化成正相关, 并且行为上的评估也表现出了过度泛化的现象.由此可见, 海马DG和CA3部分的损伤以及海马体积的减少可能是造成焦虑个体过度泛化的原因之一. ...
... 脑岛和海马、杏仁核、水管周灰质(periaqueductal gray )共同控制自主神经系统, 它们在加工恐惧情绪时被激活(Saper, 1982).前人研究表明, 当个体面对不确定的情景及对恐惧刺激进行预期时, 脑岛的响应随之增强(Berns et al., 2006; Dunsmoor, Bandettini, & Knight, 2007).一般认为, 前脑岛是躯体感觉和内脏感觉的结合点, 与ACC、杏仁核、辅助运动区(supplementary motor area, SMA)等区域一起引发与恐惧相关的行为.Dunsmoor, Prince等(2011)首次用功能成像技术结合情绪面孔感知维度对人类恐惧泛化进行研究, 并证明了在条件性恐惧的习得和表达中, 当GS接近CS时, 脑岛、尾状核(caudate)、丘脑的激活程度普遍增加.例如Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013a)在用几何图形测试人类恐惧泛化的过程中, 发现在接近CS时, 脑岛、尾状核、ACC、SMA的响应会随着刺激相似性的增加而增强.随后, Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013b)用相同的范式对广泛性焦虑障碍患者进行恐惧泛化测试, 发现前脑岛(anterior insular, AI)与ACC、杏仁核、SMA连接增强.同时, Lissek, Bradford 等(2014)用圆圈刺激发现在恐惧泛化任务中, 正常被试脑岛激活增强.此后, Onat和Büchel (2015)用面孔范式发现脑岛对CS+最敏感, 颞下皮层(inferotemporal cortex, ITC)对CS+和CS-中间的模糊刺激比较敏感, 这表明前脑岛皮层编码“危险识别”过程, 颞下皮层则对“模糊不清”的信息进行编码.可见, 脑岛在知觉的恐惧泛化过程中参与恐惧的表达和刺激的辨别. ...
... 内侧前额叶皮层(medial prefrontal cortex, mPFC)是恐惧神经环路的关键脑区, 在恐惧消退过程中起着抑制恐惧的作用(Milad et al., 2007; Orsini & Maren, 2012).近来研究表明, mPFC与恐惧泛化过程也有密切的联系.Cullen, Gilman, Winiecki, Riccio和Jasnow (2015)发现, 情景泛化记忆的表达与前扣带皮层(anterior cingulate cortex, ACC)、下边缘皮层(infralimbic cortex, IL)、前边缘皮层(prelimbic cortex, PL)这些前额叶皮层脑区的活性有关, 其中ACC与海马CA1区的连接抑制将减弱恐惧的泛化.另外, 与脑岛等脑区相反的是, vmPFC在接近CS-时激活更强(Greenberg et al., 2013a, 2013b; Lissek, Brandford et al., 2014), 在安全线索条件下GAD患者的vmPFC激活较正常人弱(Greenberg et al., 2013b).此外, vmPFC皮质厚度和恐惧泛化梯度有关, vmPFC皮质的减少可能是导致GAD患者过度泛化的原因之一(Cha et al, 2014).Xu等(2012)观察到mPFC处突触传递的毁损将导致动物情景恐惧记忆的过度泛化.他们认为mPFC、海马以及连接核(nucleus reuniens, NR) (丘脑中线核群的一部分)共同控制着恐惧记忆的泛化.具体而言, 由于mPFC是参与动机及情绪状态的关键脑区, 因此mPFC-NR路径将记忆的动机以及情绪效价信息传递至海马进行特征的重叠编码, 海马又将编码后的信息返回至mPFC进而对泛化记忆进行调节(Xu & Südhof, 2013).另有研究表明, 老鼠对声音习得的恐惧反应和mPFC处胆碱能系统(nitrergic system) (神经元轴突分泌的一类神经递质)的激活水平能够预测泛化的程度, 表现为胆碱能系统激活程度越高, 泛化程度越高(Saul’skaya & Sudorgina, 2016).此外, Likhtik, Stujenske, Topiwala, Harris和Gordon (2014)认为内侧前额叶(mPFC)以及基底外侧核(BLA)连接的增强预示着内侧前额叶可能会影响基底外侧核的活动, 并通过抑制杏仁核的活动来减弱恐惧反应.相反的, mPFC-BLA连接的减弱则会导致对杏仁核的抑制失败, 从而增强恐惧反应.可见, mPFC在知觉恐惧泛化过程中参与调节和抑制恐惧. ...
Generalized anxiety disorder is associated with overgeneralization of classically conditioned fear
3
2014
... 恐惧是一种对环境的适应性机制, 当危险发生时, 恐惧使得机体启动防御机制并对危险刺激进行反应(Dymond, Dunsmoor, Vervliet, Roche, & Hermans, 2015; Hoppenbrouwers, Bulten, & Brazil, 2016; LeDoux, 2000).人类的恐惧反应多是后天习得, 例如, 在华生的条件反射实验中, 小阿尔伯特原本并不惧怕白鼠, 然而当白鼠与巨大的金属响声(危险刺激)同时出现后, 小阿尔伯特便对白鼠产生了恐惧反应, 此后当他再见到小白兔、白色绒毛玩具、甚至老人的白胡须等相关事物时, 他都会感到害怕(Watson & Rayner, 1920).这种不仅对特定的刺激(白鼠)产生恐惧, 而且对与特定刺激相关的事物同样会产生恐惧的现象即是“恐惧泛化” (Dunsmoor, Prince, Murty, Kragel, & LaBar, 2011; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014; Onat & Büchel, 2015).一般来说, 适当的恐惧泛化对个体生存具有积极意义, 因为在漫长的进化过程中, 恐惧泛化能够促使人类躲避各种潜在威胁, 有利于个体生存繁衍; 而在日常生活中, 恐惧泛化也可增强个体的警觉性, 从而有效并快速地对危险环境做出预测和应对(Steimer, 2002). ...
... 大量恐惧泛化的研究表明, 相同知觉维度的刺激可以根据物理上的相似性由高到低被分成几个不同的层次, 而个体的恐惧反应也随着刺激的相似性高低而呈现出不同反应大小的梯度 (Ghirlanda & Enquist, 2003; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014).在知觉恐惧泛化研究中, 由一个感觉刺激(记为CS+; 如特定的灯光、声音、形状)与恐惧刺激(US; 如电击、恐惧图片、尖叫声)建立的条件反应, 另一个区别于CS+的感觉刺激作为对照刺激从不匹配US (记为CS-; 如与CS+差别较大的灯光、声音、形状), 通常用相同知觉维度的刺激(记为GS; 如在CS+和CS-之间变化的灯光、声音、形状)进行泛化测试(Hermans, Baeyens, & Vervliet., 2013).这种基于知觉的恐惧泛化梯度可以反映辨别学习和恐惧的泛化程度(Lissek et al., 2008; Struyf, Zaman, Vervliet, & van Diest, 2015) (见图1). ...
... 图形特征在知觉恐惧泛化研究中被广泛运用, 主要包括图形大小、颜色、形状等特征的变化.首先, 在图形大小方面, Lissek等(2008)曾用一大一小两个圆圈分别作为CS+和CS-, 8个大小处在CS+和CS-之间的圆圈作为GS测试被试的恐惧反应, 结果发现被试的眨眼惊跳反应随着GS与CS+大小的相似性增加而增强.随后, 研究者用该范式进一步确认, 广泛性焦虑障碍和惊恐障碍患者的恐惧泛化比正常被试更广(Lissek et al., 2010; Lissek, Kaczkurkin et al., 2014).近期, 使用该范式的国内研究发现, 状态焦虑会增加恐惧的泛化, 而积极情绪会减弱恐惧的泛化(徐亮等, 2016; 冯彪等, 2017). ...
Overgeneralization of conditioned fear as a pathogenic marker of panic disorder
3
2010
... 恐惧是一种对环境的适应性机制, 当危险发生时, 恐惧使得机体启动防御机制并对危险刺激进行反应(Dymond, Dunsmoor, Vervliet, Roche, & Hermans, 2015; Hoppenbrouwers, Bulten, & Brazil, 2016; LeDoux, 2000).人类的恐惧反应多是后天习得, 例如, 在华生的条件反射实验中, 小阿尔伯特原本并不惧怕白鼠, 然而当白鼠与巨大的金属响声(危险刺激)同时出现后, 小阿尔伯特便对白鼠产生了恐惧反应, 此后当他再见到小白兔、白色绒毛玩具、甚至老人的白胡须等相关事物时, 他都会感到害怕(Watson & Rayner, 1920).这种不仅对特定的刺激(白鼠)产生恐惧, 而且对与特定刺激相关的事物同样会产生恐惧的现象即是“恐惧泛化” (Dunsmoor, Prince, Murty, Kragel, & LaBar, 2011; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014; Onat & Büchel, 2015).一般来说, 适当的恐惧泛化对个体生存具有积极意义, 因为在漫长的进化过程中, 恐惧泛化能够促使人类躲避各种潜在威胁, 有利于个体生存繁衍; 而在日常生活中, 恐惧泛化也可增强个体的警觉性, 从而有效并快速地对危险环境做出预测和应对(Steimer, 2002). ...
... 大量恐惧泛化的研究表明, 相同知觉维度的刺激可以根据物理上的相似性由高到低被分成几个不同的层次, 而个体的恐惧反应也随着刺激的相似性高低而呈现出不同反应大小的梯度 (Ghirlanda & Enquist, 2003; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014).在知觉恐惧泛化研究中, 由一个感觉刺激(记为CS+; 如特定的灯光、声音、形状)与恐惧刺激(US; 如电击、恐惧图片、尖叫声)建立的条件反应, 另一个区别于CS+的感觉刺激作为对照刺激从不匹配US (记为CS-; 如与CS+差别较大的灯光、声音、形状), 通常用相同知觉维度的刺激(记为GS; 如在CS+和CS-之间变化的灯光、声音、形状)进行泛化测试(Hermans, Baeyens, & Vervliet., 2013).这种基于知觉的恐惧泛化梯度可以反映辨别学习和恐惧的泛化程度(Lissek et al., 2008; Struyf, Zaman, Vervliet, & van Diest, 2015) (见图1). ...
... 图形特征在知觉恐惧泛化研究中被广泛运用, 主要包括图形大小、颜色、形状等特征的变化.首先, 在图形大小方面, Lissek等(2008)曾用一大一小两个圆圈分别作为CS+和CS-, 8个大小处在CS+和CS-之间的圆圈作为GS测试被试的恐惧反应, 结果发现被试的眨眼惊跳反应随着GS与CS+大小的相似性增加而增强.随后, 研究者用该范式进一步确认, 广泛性焦虑障碍和惊恐障碍患者的恐惧泛化比正常被试更广(Lissek et al., 2010; Lissek, Kaczkurkin et al., 2014).近期, 使用该范式的国内研究发现, 状态焦虑会增加恐惧的泛化, 而积极情绪会减弱恐惧的泛化(徐亮等, 2016; 冯彪等, 2017). ...
Neuroticism and avoidance of ambiguous stimuli: Better safe than sorry?
1
2010
... 图形颜色特征变化也很常见.与大小变化相似, 研究者通常采用两个颜色差别明显的圆圈分别作为CS+和CS-, 再用颜色处在之间变化的圆圈测试被试的恐惧反应.例如Lommen, Engelhard和van den Hout (2010)采用白色(CS+)、黑色(CS-)和灰色(GS)圆圈作为实验材料, 发现神经质特质人群的恐惧泛化存在着显著差异, 表现为高神经质组在面对模棱两可的刺激时表现出比低神经质组更多的恐惧反应.Dunsmoor和LaBar (2013)通过操纵CS-的颜色来探讨CS-特征对恐惧泛化的影响, 即用蓝绿色圆圈作为CS+, 绿色或蓝色圆圈作为CS-, 处在CS+和CS-之间的变化的圆圈测试恐惧泛化反应, 结果发现被试对远离CS-的刺激产生了泛化. ...
Neural circuits and mechanisms involved in fear generalization: Implications for the pathophysiology and treatment of posttraumatic stress disorder
1
2016
... 杏仁核是恐惧学习和恐惧泛化的关键脑区, 是CS与US的汇聚点并启动由威胁引起的条件反应.运用情景辨别任务, Ciocchi等(2010)发现激活中央杏仁核外侧核(lateral part of central amygdala, CeL)神经元的活动, 或者抑制中央杏仁核内侧核(medial part of central amygdala, CeM)神经元的活动都会增加老鼠对CS-的恐惧反应.这说明杏仁核中央核团(central amygdala, CeA)的神经活性对恐惧泛化具有一定的调节作用.Lopresto, Schipper和Homberg (2016)认为探索杏仁核在恐惧泛化中的作用即是把杏仁核看作神经回路的节点, 该节点连接着海马和大脑皮层.在情景任务中, 已有研究表明恐惧情景引发老鼠和人类的杏仁核以及海马的响应比中性情景更强(Maren, Phan, & Liberzon, 2013; Marschner, Kalisch, Vervliet, Vansteenwegen, & Buchel, 2008).这说明, 杏仁核以及海马可能是调节情景恐惧泛化的关键脑区之一.此后, Resnik和Paz (2015)用纯音作为刺激材料考察了猴子恐惧泛化的神经机制及其行为反应, 发现基底外侧核(basolateral complex of the amygdala, BLA)处神经元的特异变化减弱了泛化阶段的辨别学习, 这种杏仁核部位的轻微改变影响了前额叶皮层以及海马, 并破坏了泛化的平衡, 从而导致了更广的泛化.另外, 内侧前额叶(mPFC)-BLA通路的增强可以促进恐惧消退, 这种自上而下连接不良的传递, 也会导致更广的泛化(Jasnow et al., 2017; Likhtik & Paz, 2015).由此可见, 杏仁核在知觉恐惧泛化回路中有着重要的作用, 是其他脑区与感知觉信息的连接点, 控制着恐惧反应的表达. ...
Sex differences in the generalization of fear as a function of retention intervals
1
2013
... 情景的恐惧泛化需要对整个情景内的信息进行辨别, 这些信息通常包括空间、嗅觉、听觉和视觉信息, 而不是单一的物体特征的变化(Jasnow, Lynch, Gilman, & Riccio, 2017).首先, 在动物研究方面, 有研究者训练啮齿类动物对某一情景(CXT+)习得恐惧, 而在另外一个新异的情景(CXT-)下始终不匹配恐惧刺激, 紧接着在一个和CXT+相似的情景(G-CXT)中进行恐惧泛化测试.这些情景任务包括辨别触摸屏中物体与屏幕的距离(Mcallister, Saksida, & Bussey, 2013; Talpos, McTighe, Dias, Saksida, & Bussey, 2010)、辨别新异刺激和熟悉刺激的位置(Bekinschtein et al., 2013)、在不同形状的空间里习得恐惧(Mchugh et al., 2007)等.动物研究表明, 恐惧习得和重现刺激情景(G-CXT)之间的时间间隔越长, 恐惧泛化程度越大(Biedenkapp & Rudy, 2007).而在习得任务之前预先暴露情景则会减弱恐惧的泛化.此外, 研究者通过运用情景恐惧泛化范式, 发现啮齿动物恐惧泛化存在着性别差异, 具体表现为雌性比雄性更容易发生泛化(Keiser et al., 2017), 且泛化速度更快(Lynch, Cullen, Jasnow, & Riccio, 2013).预先暴露情景增加了雄性老鼠在习得阶段的恐惧反应, 但却减少了雌性老鼠在泛化的阶段恐惧反应(Keiser et al., 2017). ...
Activation of ERβ modulates fear generalization through an effect on memory retrieval
1
2014
... 鉴于恐惧泛化几乎是所有焦虑障碍的主要症状, 而女性又是焦虑障碍的易感人群 (McLean, Asnaani, Litz, & Hofmann, 2011), 因此分析女性激素对恐惧泛化的影响具有重要的临床意义.研究表明, 恐惧泛化的性别差异很可能是由雌二醇导致(Lynch et al., 2014), 雌二醇或通过促进海马的“模式完成”或者损害“模式分离”来扩大恐惧泛化(Lynch, Winiecki, Vanderhoof, Riccio, & Jasnow, 2016).此外, 催产素有减缓焦虑情绪的作用, 因此可能对治疗焦虑症有重要价值.近期科学家甚至发现催产素可以降低人类和动物的恐惧反应, 促进恐惧的消退(Eckstein et al., 2015; Hyun, Perry, Ganella, & Madsen, 2017; Missig, Ayers, Schulkin, & Rosen, 2010).如把催产素注射到啮齿动物的中央杏仁核(central amygdala)或者背外侧隔膜(dorso-lateral septum)可以减少在消退阶段的恐惧反应(Viviani & Stoop, 2008; Zoicas, Slattery, & Neumann, 2014).在对人类进行的研究中, 使用鼻喷催产素后, 被试mPFC的活动明显增强从而可以达到促使恐惧消退的效果. ...
Hippocampal cytosolic estrogen receptors regulate fear generalization in females
1
2016
... 鉴于恐惧泛化几乎是所有焦虑障碍的主要症状, 而女性又是焦虑障碍的易感人群 (McLean, Asnaani, Litz, & Hofmann, 2011), 因此分析女性激素对恐惧泛化的影响具有重要的临床意义.研究表明, 恐惧泛化的性别差异很可能是由雌二醇导致(Lynch et al., 2014), 雌二醇或通过促进海马的“模式完成”或者损害“模式分离”来扩大恐惧泛化(Lynch, Winiecki, Vanderhoof, Riccio, & Jasnow, 2016).此外, 催产素有减缓焦虑情绪的作用, 因此可能对治疗焦虑症有重要价值.近期科学家甚至发现催产素可以降低人类和动物的恐惧反应, 促进恐惧的消退(Eckstein et al., 2015; Hyun, Perry, Ganella, & Madsen, 2017; Missig, Ayers, Schulkin, & Rosen, 2010).如把催产素注射到啮齿动物的中央杏仁核(central amygdala)或者背外侧隔膜(dorso-lateral septum)可以减少在消退阶段的恐惧反应(Viviani & Stoop, 2008; Zoicas, Slattery, & Neumann, 2014).在对人类进行的研究中, 使用鼻喷催产素后, 被试mPFC的活动明显增强从而可以达到促使恐惧消退的效果. ...
The contextual brain: Implications for fear conditioning, extinction and psychopathology
1
2013
... 杏仁核是恐惧学习和恐惧泛化的关键脑区, 是CS与US的汇聚点并启动由威胁引起的条件反应.运用情景辨别任务, Ciocchi等(2010)发现激活中央杏仁核外侧核(lateral part of central amygdala, CeL)神经元的活动, 或者抑制中央杏仁核内侧核(medial part of central amygdala, CeM)神经元的活动都会增加老鼠对CS-的恐惧反应.这说明杏仁核中央核团(central amygdala, CeA)的神经活性对恐惧泛化具有一定的调节作用.Lopresto, Schipper和Homberg (2016)认为探索杏仁核在恐惧泛化中的作用即是把杏仁核看作神经回路的节点, 该节点连接着海马和大脑皮层.在情景任务中, 已有研究表明恐惧情景引发老鼠和人类的杏仁核以及海马的响应比中性情景更强(Maren, Phan, & Liberzon, 2013; Marschner, Kalisch, Vervliet, Vansteenwegen, & Buchel, 2008).这说明, 杏仁核以及海马可能是调节情景恐惧泛化的关键脑区之一.此后, Resnik和Paz (2015)用纯音作为刺激材料考察了猴子恐惧泛化的神经机制及其行为反应, 发现基底外侧核(basolateral complex of the amygdala, BLA)处神经元的特异变化减弱了泛化阶段的辨别学习, 这种杏仁核部位的轻微改变影响了前额叶皮层以及海马, 并破坏了泛化的平衡, 从而导致了更广的泛化.另外, 内侧前额叶(mPFC)-BLA通路的增强可以促进恐惧消退, 这种自上而下连接不良的传递, 也会导致更广的泛化(Jasnow et al., 2017; Likhtik & Paz, 2015).由此可见, 杏仁核在知觉恐惧泛化回路中有着重要的作用, 是其他脑区与感知觉信息的连接点, 控制着恐惧反应的表达. ...
Dissociable roles for the hippocampus and the amygdala in human cued versus context fear conditioning
1
2008
... 杏仁核是恐惧学习和恐惧泛化的关键脑区, 是CS与US的汇聚点并启动由威胁引起的条件反应.运用情景辨别任务, Ciocchi等(2010)发现激活中央杏仁核外侧核(lateral part of central amygdala, CeL)神经元的活动, 或者抑制中央杏仁核内侧核(medial part of central amygdala, CeM)神经元的活动都会增加老鼠对CS-的恐惧反应.这说明杏仁核中央核团(central amygdala, CeA)的神经活性对恐惧泛化具有一定的调节作用.Lopresto, Schipper和Homberg (2016)认为探索杏仁核在恐惧泛化中的作用即是把杏仁核看作神经回路的节点, 该节点连接着海马和大脑皮层.在情景任务中, 已有研究表明恐惧情景引发老鼠和人类的杏仁核以及海马的响应比中性情景更强(Maren, Phan, & Liberzon, 2013; Marschner, Kalisch, Vervliet, Vansteenwegen, & Buchel, 2008).这说明, 杏仁核以及海马可能是调节情景恐惧泛化的关键脑区之一.此后, Resnik和Paz (2015)用纯音作为刺激材料考察了猴子恐惧泛化的神经机制及其行为反应, 发现基底外侧核(basolateral complex of the amygdala, BLA)处神经元的特异变化减弱了泛化阶段的辨别学习, 这种杏仁核部位的轻微改变影响了前额叶皮层以及海马, 并破坏了泛化的平衡, 从而导致了更广的泛化.另外, 内侧前额叶(mPFC)-BLA通路的增强可以促进恐惧消退, 这种自上而下连接不良的传递, 也会导致更广的泛化(Jasnow et al., 2017; Likhtik & Paz, 2015).由此可见, 杏仁核在知觉恐惧泛化回路中有着重要的作用, 是其他脑区与感知觉信息的连接点, 控制着恐惧反应的表达. ...
Dissociation between memory retention across a delay and pattern separation following medial prefrontal cortex lesions in the touchscreen TUNL task
1
2013
... 情景的恐惧泛化需要对整个情景内的信息进行辨别, 这些信息通常包括空间、嗅觉、听觉和视觉信息, 而不是单一的物体特征的变化(Jasnow, Lynch, Gilman, & Riccio, 2017).首先, 在动物研究方面, 有研究者训练啮齿类动物对某一情景(CXT+)习得恐惧, 而在另外一个新异的情景(CXT-)下始终不匹配恐惧刺激, 紧接着在一个和CXT+相似的情景(G-CXT)中进行恐惧泛化测试.这些情景任务包括辨别触摸屏中物体与屏幕的距离(Mcallister, Saksida, & Bussey, 2013; Talpos, McTighe, Dias, Saksida, & Bussey, 2010)、辨别新异刺激和熟悉刺激的位置(Bekinschtein et al., 2013)、在不同形状的空间里习得恐惧(Mchugh et al., 2007)等.动物研究表明, 恐惧习得和重现刺激情景(G-CXT)之间的时间间隔越长, 恐惧泛化程度越大(Biedenkapp & Rudy, 2007).而在习得任务之前预先暴露情景则会减弱恐惧的泛化.此外, 研究者通过运用情景恐惧泛化范式, 发现啮齿动物恐惧泛化存在着性别差异, 具体表现为雌性比雄性更容易发生泛化(Keiser et al., 2017), 且泛化速度更快(Lynch, Cullen, Jasnow, & Riccio, 2013).预先暴露情景增加了雄性老鼠在习得阶段的恐惧反应, 但却减少了雌性老鼠在泛化的阶段恐惧反应(Keiser et al., 2017). ...
Dentate gyrus NMDA receptors mediate rapid pattern separation in the hippocampal network
1
2007
... 情景的恐惧泛化需要对整个情景内的信息进行辨别, 这些信息通常包括空间、嗅觉、听觉和视觉信息, 而不是单一的物体特征的变化(Jasnow, Lynch, Gilman, & Riccio, 2017).首先, 在动物研究方面, 有研究者训练啮齿类动物对某一情景(CXT+)习得恐惧, 而在另外一个新异的情景(CXT-)下始终不匹配恐惧刺激, 紧接着在一个和CXT+相似的情景(G-CXT)中进行恐惧泛化测试.这些情景任务包括辨别触摸屏中物体与屏幕的距离(Mcallister, Saksida, & Bussey, 2013; Talpos, McTighe, Dias, Saksida, & Bussey, 2010)、辨别新异刺激和熟悉刺激的位置(Bekinschtein et al., 2013)、在不同形状的空间里习得恐惧(Mchugh et al., 2007)等.动物研究表明, 恐惧习得和重现刺激情景(G-CXT)之间的时间间隔越长, 恐惧泛化程度越大(Biedenkapp & Rudy, 2007).而在习得任务之前预先暴露情景则会减弱恐惧的泛化.此外, 研究者通过运用情景恐惧泛化范式, 发现啮齿动物恐惧泛化存在着性别差异, 具体表现为雌性比雄性更容易发生泛化(Keiser et al., 2017), 且泛化速度更快(Lynch, Cullen, Jasnow, & Riccio, 2013).预先暴露情景增加了雄性老鼠在习得阶段的恐惧反应, 但却减少了雌性老鼠在泛化的阶段恐惧反应(Keiser et al., 2017). ...
Gender differences in anxiety disorders: Prevalence, course of illness, comorbidity and burden of illness
1
2011
... 鉴于恐惧泛化几乎是所有焦虑障碍的主要症状, 而女性又是焦虑障碍的易感人群 (McLean, Asnaani, Litz, & Hofmann, 2011), 因此分析女性激素对恐惧泛化的影响具有重要的临床意义.研究表明, 恐惧泛化的性别差异很可能是由雌二醇导致(Lynch et al., 2014), 雌二醇或通过促进海马的“模式完成”或者损害“模式分离”来扩大恐惧泛化(Lynch, Winiecki, Vanderhoof, Riccio, & Jasnow, 2016).此外, 催产素有减缓焦虑情绪的作用, 因此可能对治疗焦虑症有重要价值.近期科学家甚至发现催产素可以降低人类和动物的恐惧反应, 促进恐惧的消退(Eckstein et al., 2015; Hyun, Perry, Ganella, & Madsen, 2017; Missig, Ayers, Schulkin, & Rosen, 2010).如把催产素注射到啮齿动物的中央杏仁核(central amygdala)或者背外侧隔膜(dorso-lateral septum)可以减少在消退阶段的恐惧反应(Viviani & Stoop, 2008; Zoicas, Slattery, & Neumann, 2014).在对人类进行的研究中, 使用鼻喷催产素后, 被试mPFC的活动明显增强从而可以达到促使恐惧消退的效果. ...
Recall of fear extinction in humans activates the ventromedial prefrontal cortex and hippocampus in concert
2
2007
... 巴普洛夫经典条件反射理论认为, 当安全的条件刺激(conditioned stimulus, CS)和一个令人恐惧的无条件刺激(unconditioned stimulus, US)多次匹配(CS-US)以后, 单独呈现条件刺激即能引起人出汗、呼吸加快、心跳加速等条件性恐惧反应(conditioned responses, CR).巴普洛夫经典条件反射已被广泛应用于研究恐惧的习得、表达、消退的研究中(Lang, Davis, & Öhman, 2000; LeDoux, 1996; Milad et al., 2007).然而恐惧学习并不仅限于恐惧条件化情形之下的特定事件或事物, 任何与恐惧刺激相关的事物都有可能引发恐惧反应, 这即是恐惧泛化(fear generalization), 而那些从未与无条件刺激匹配却能够引起恐惧反应的相关刺激便被称为泛化刺激(generalization stimulus, GS). ...
... 内侧前额叶皮层(medial prefrontal cortex, mPFC)是恐惧神经环路的关键脑区, 在恐惧消退过程中起着抑制恐惧的作用(Milad et al., 2007; Orsini & Maren, 2012).近来研究表明, mPFC与恐惧泛化过程也有密切的联系.Cullen, Gilman, Winiecki, Riccio和Jasnow (2015)发现, 情景泛化记忆的表达与前扣带皮层(anterior cingulate cortex, ACC)、下边缘皮层(infralimbic cortex, IL)、前边缘皮层(prelimbic cortex, PL)这些前额叶皮层脑区的活性有关, 其中ACC与海马CA1区的连接抑制将减弱恐惧的泛化.另外, 与脑岛等脑区相反的是, vmPFC在接近CS-时激活更强(Greenberg et al., 2013a, 2013b; Lissek, Brandford et al., 2014), 在安全线索条件下GAD患者的vmPFC激活较正常人弱(Greenberg et al., 2013b).此外, vmPFC皮质厚度和恐惧泛化梯度有关, vmPFC皮质的减少可能是导致GAD患者过度泛化的原因之一(Cha et al, 2014).Xu等(2012)观察到mPFC处突触传递的毁损将导致动物情景恐惧记忆的过度泛化.他们认为mPFC、海马以及连接核(nucleus reuniens, NR) (丘脑中线核群的一部分)共同控制着恐惧记忆的泛化.具体而言, 由于mPFC是参与动机及情绪状态的关键脑区, 因此mPFC-NR路径将记忆的动机以及情绪效价信息传递至海马进行特征的重叠编码, 海马又将编码后的信息返回至mPFC进而对泛化记忆进行调节(Xu & Südhof, 2013).另有研究表明, 老鼠对声音习得的恐惧反应和mPFC处胆碱能系统(nitrergic system) (神经元轴突分泌的一类神经递质)的激活水平能够预测泛化的程度, 表现为胆碱能系统激活程度越高, 泛化程度越高(Saul’skaya & Sudorgina, 2016).此外, Likhtik, Stujenske, Topiwala, Harris和Gordon (2014)认为内侧前额叶(mPFC)以及基底外侧核(BLA)连接的增强预示着内侧前额叶可能会影响基底外侧核的活动, 并通过抑制杏仁核的活动来减弱恐惧反应.相反的, mPFC-BLA连接的减弱则会导致对杏仁核的抑制失败, 从而增强恐惧反应.可见, mPFC在知觉恐惧泛化过程中参与调节和抑制恐惧. ...
Oxytocin reduces background anxiety in a fear-potentiated startle paradigm
1
2010
... 鉴于恐惧泛化几乎是所有焦虑障碍的主要症状, 而女性又是焦虑障碍的易感人群 (McLean, Asnaani, Litz, & Hofmann, 2011), 因此分析女性激素对恐惧泛化的影响具有重要的临床意义.研究表明, 恐惧泛化的性别差异很可能是由雌二醇导致(Lynch et al., 2014), 雌二醇或通过促进海马的“模式完成”或者损害“模式分离”来扩大恐惧泛化(Lynch, Winiecki, Vanderhoof, Riccio, & Jasnow, 2016).此外, 催产素有减缓焦虑情绪的作用, 因此可能对治疗焦虑症有重要价值.近期科学家甚至发现催产素可以降低人类和动物的恐惧反应, 促进恐惧的消退(Eckstein et al., 2015; Hyun, Perry, Ganella, & Madsen, 2017; Missig, Ayers, Schulkin, & Rosen, 2010).如把催产素注射到啮齿动物的中央杏仁核(central amygdala)或者背外侧隔膜(dorso-lateral septum)可以减少在消退阶段的恐惧反应(Viviani & Stoop, 2008; Zoicas, Slattery, & Neumann, 2014).在对人类进行的研究中, 使用鼻喷催产素后, 被试mPFC的活动明显增强从而可以达到促使恐惧消退的效果. ...
Suppression of adult neurogenesis impairs population coding of similar contexts in hippocampal CA3 region
1
2012
... 海马中 “模式分离(pattern separation)”和“模式完成(pattern completion)”负责对刺激的属性特征与已存储的信息进行匹配(Gluck & Myers, 1993).“模式分离”由海马的齿状回执行(dentate gyrus, DG), 是一种把相似信息区分开的神经活动; 而“模式完成”主要发生在海马CA3区, 它描述了相反的加工过程, 如果刺激的区别较小, “模式完成”则会忽略刺激间的区别(Drew & Huckleberry, 2017; Gilbert, Kesner, & Lee, 2001; Treves & Rolls, 1992).在知觉泛化过程中, 当GS和CS相似性较高时, 海马启动“模式完成”使CS的神经表征激活, 从而进一步引发被试的恐惧反应; 而当GS和CS相似性较低时, 海马则启动“模式分离”激活腹内测前额叶从而降低杏仁核的活动, 达到抑制恐惧反应的效果(Lissek, 2012; Lissek, Bradford et al., 2014).例如, 研究发现在啮齿动物的情景辨别恐惧泛化任务中, 海马CA3区的神经抑制会促进啮齿动物在恐惧和中性情景下的“模式完成”过程, 从而增加恐惧的泛化; 反过来, 齿状回处的神经兴奋则会促进“模式分离”过程, 以抑制恐惧泛化(Sahay et al., 2011; Niibori et al., 2012).可见, 海马在知觉恐惧泛化的过程中负责刺激的辨别加工. ...
The neuronal basis of fear generalization in humans
4
2015
... 恐惧是一种对环境的适应性机制, 当危险发生时, 恐惧使得机体启动防御机制并对危险刺激进行反应(Dymond, Dunsmoor, Vervliet, Roche, & Hermans, 2015; Hoppenbrouwers, Bulten, & Brazil, 2016; LeDoux, 2000).人类的恐惧反应多是后天习得, 例如, 在华生的条件反射实验中, 小阿尔伯特原本并不惧怕白鼠, 然而当白鼠与巨大的金属响声(危险刺激)同时出现后, 小阿尔伯特便对白鼠产生了恐惧反应, 此后当他再见到小白兔、白色绒毛玩具、甚至老人的白胡须等相关事物时, 他都会感到害怕(Watson & Rayner, 1920).这种不仅对特定的刺激(白鼠)产生恐惧, 而且对与特定刺激相关的事物同样会产生恐惧的现象即是“恐惧泛化” (Dunsmoor, Prince, Murty, Kragel, & LaBar, 2011; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014; Onat & Büchel, 2015).一般来说, 适当的恐惧泛化对个体生存具有积极意义, 因为在漫长的进化过程中, 恐惧泛化能够促使人类躲避各种潜在威胁, 有利于个体生存繁衍; 而在日常生活中, 恐惧泛化也可增强个体的警觉性, 从而有效并快速地对危险环境做出预测和应对(Steimer, 2002). ...
... 然而, 过度的恐惧泛化则会对个体生存产生不利影响.一方面, 过度的恐惧泛化被认为是广泛性焦虑障碍(generalized anxiety disorder, GAD)、创伤后应激障碍(post-traumatic stress disorder, PTSD)、惊恐障碍(panic disorder, PD)、特殊恐惧症(Specific phobia)等焦虑障碍的重要致病因素之一(Onat & Büchel, 2015), 如个体遭受严重车祸后, 在面对与车祸相关的线索或者情景(如红绿灯路口、鸣笛声等)时则会表现出恐惧情绪及回避反应; 另一方面, 恐惧泛化不仅仅限于外在刺激, 如果个体在经历创伤性事件时有心跳加速的症状, 那么与此相联系的其他内在感觉, 如胸闷、头昏的症状也可能引发恐惧(Bouton, Mineka, & Barlow, 2001).由此可见, 过度的恐惧泛化将会导致个体过度焦虑、社会功能受损和生活质量下降等诸多问题, 从而给患者的正常生活造成巨大困扰.因此, 恐惧泛化研究对克服焦虑障碍具有重要的临床意义. ...
... 第一类研究通常以两张同性别个体的中性面孔作为CS+和CS-, GS则是在CS+和CS-之间融合的面孔.如, Onat和Büchel (2015)运用该范式发现被试的皮肤电水平随着面孔的相似性增加而增强.另外, Holt等(2014)采用该法并结合差别域限法(JND)设置GS.GS被分成能够察觉的面孔和不能察觉的面孔两大类.结果表明, 阈限下的面孔比阈限上的面孔能引发更大的恐惧反应. ...
... 脑岛和海马、杏仁核、水管周灰质(periaqueductal gray )共同控制自主神经系统, 它们在加工恐惧情绪时被激活(Saper, 1982).前人研究表明, 当个体面对不确定的情景及对恐惧刺激进行预期时, 脑岛的响应随之增强(Berns et al., 2006; Dunsmoor, Bandettini, & Knight, 2007).一般认为, 前脑岛是躯体感觉和内脏感觉的结合点, 与ACC、杏仁核、辅助运动区(supplementary motor area, SMA)等区域一起引发与恐惧相关的行为.Dunsmoor, Prince等(2011)首次用功能成像技术结合情绪面孔感知维度对人类恐惧泛化进行研究, 并证明了在条件性恐惧的习得和表达中, 当GS接近CS时, 脑岛、尾状核(caudate)、丘脑的激活程度普遍增加.例如Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013a)在用几何图形测试人类恐惧泛化的过程中, 发现在接近CS时, 脑岛、尾状核、ACC、SMA的响应会随着刺激相似性的增加而增强.随后, Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013b)用相同的范式对广泛性焦虑障碍患者进行恐惧泛化测试, 发现前脑岛(anterior insular, AI)与ACC、杏仁核、SMA连接增强.同时, Lissek, Bradford 等(2014)用圆圈刺激发现在恐惧泛化任务中, 正常被试脑岛激活增强.此后, Onat和Büchel (2015)用面孔范式发现脑岛对CS+最敏感, 颞下皮层(inferotemporal cortex, ITC)对CS+和CS-中间的模糊刺激比较敏感, 这表明前脑岛皮层编码“危险识别”过程, 颞下皮层则对“模糊不清”的信息进行编码.可见, 脑岛在知觉的恐惧泛化过程中参与恐惧的表达和刺激的辨别. ...
Neural and cellular mechanisms of fear and extinction memory formation
1
2012
... 内侧前额叶皮层(medial prefrontal cortex, mPFC)是恐惧神经环路的关键脑区, 在恐惧消退过程中起着抑制恐惧的作用(Milad et al., 2007; Orsini & Maren, 2012).近来研究表明, mPFC与恐惧泛化过程也有密切的联系.Cullen, Gilman, Winiecki, Riccio和Jasnow (2015)发现, 情景泛化记忆的表达与前扣带皮层(anterior cingulate cortex, ACC)、下边缘皮层(infralimbic cortex, IL)、前边缘皮层(prelimbic cortex, PL)这些前额叶皮层脑区的活性有关, 其中ACC与海马CA1区的连接抑制将减弱恐惧的泛化.另外, 与脑岛等脑区相反的是, vmPFC在接近CS-时激活更强(Greenberg et al., 2013a, 2013b; Lissek, Brandford et al., 2014), 在安全线索条件下GAD患者的vmPFC激活较正常人弱(Greenberg et al., 2013b).此外, vmPFC皮质厚度和恐惧泛化梯度有关, vmPFC皮质的减少可能是导致GAD患者过度泛化的原因之一(Cha et al, 2014).Xu等(2012)观察到mPFC处突触传递的毁损将导致动物情景恐惧记忆的过度泛化.他们认为mPFC、海马以及连接核(nucleus reuniens, NR) (丘脑中线核群的一部分)共同控制着恐惧记忆的泛化.具体而言, 由于mPFC是参与动机及情绪状态的关键脑区, 因此mPFC-NR路径将记忆的动机以及情绪效价信息传递至海马进行特征的重叠编码, 海马又将编码后的信息返回至mPFC进而对泛化记忆进行调节(Xu & Südhof, 2013).另有研究表明, 老鼠对声音习得的恐惧反应和mPFC处胆碱能系统(nitrergic system) (神经元轴突分泌的一类神经递质)的激活水平能够预测泛化的程度, 表现为胆碱能系统激活程度越高, 泛化程度越高(Saul’skaya & Sudorgina, 2016).此外, Likhtik, Stujenske, Topiwala, Harris和Gordon (2014)认为内侧前额叶(mPFC)以及基底外侧核(BLA)连接的增强预示着内侧前额叶可能会影响基底外侧核的活动, 并通过抑制杏仁核的活动来减弱恐惧反应.相反的, mPFC-BLA连接的减弱则会导致对杏仁核的抑制失败, 从而增强恐惧反应.可见, mPFC在知觉恐惧泛化过程中参与调节和抑制恐惧. ...
Neuroendocrinology of post-traumatic stress disorder
1
2010
... 此外, 去甲肾上腺素(norepinephrine, NE)也已成功应用于PTSD的治疗与防治(Brunet et al., 2008; Pervanidou & Chrousos, 2010).动物实验表明, 去甲肾上腺素可能通过调节小鼠内侧前额叶皮层(mPFC)参与调节恐惧的回路, 从而对消退恐惧障碍起到改善作用(Fitzgerald, Giustino, Seemann, & Maren, 2015). ...
Fear generalization in the primate amygdala
2
2015
... 听觉是仅次于视觉的重要感觉通道, 也是动物和人类感知环境、逃避危险的线索之一.在恐惧泛化的研究中, 研究者们根据频率把声音划分成等距且可辨别的几段音频, 并与电击匹配, 来研究听觉恐惧泛化的机制.动物研究中对恐惧泛化脑机制的探索通常采用声音辨别任务(Ghosh & Chattarji, 2015; Resnik & Paz, 2015).Hovland等(1937)首次将该方法用以研究人类的恐惧泛化, 实验采用4个振幅相同、声音频率等距且可辨别的纯音作为材料, 结果发现被试对从未与US匹配的其他三个纯音也会产生恐惧反应.此外, 研究者用声音结合不同强度的电击, 探讨不同强度电击对恐惧泛化的影响, 结果表明, 不同强度的电击对被试习得恐惧并没有产生影响, 但高强度的电击却可以增加泛化的范围(Dunsmoor, Kroes, Braren, & Phelps, 2017). ...
... 杏仁核是恐惧学习和恐惧泛化的关键脑区, 是CS与US的汇聚点并启动由威胁引起的条件反应.运用情景辨别任务, Ciocchi等(2010)发现激活中央杏仁核外侧核(lateral part of central amygdala, CeL)神经元的活动, 或者抑制中央杏仁核内侧核(medial part of central amygdala, CeM)神经元的活动都会增加老鼠对CS-的恐惧反应.这说明杏仁核中央核团(central amygdala, CeA)的神经活性对恐惧泛化具有一定的调节作用.Lopresto, Schipper和Homberg (2016)认为探索杏仁核在恐惧泛化中的作用即是把杏仁核看作神经回路的节点, 该节点连接着海马和大脑皮层.在情景任务中, 已有研究表明恐惧情景引发老鼠和人类的杏仁核以及海马的响应比中性情景更强(Maren, Phan, & Liberzon, 2013; Marschner, Kalisch, Vervliet, Vansteenwegen, & Buchel, 2008).这说明, 杏仁核以及海马可能是调节情景恐惧泛化的关键脑区之一.此后, Resnik和Paz (2015)用纯音作为刺激材料考察了猴子恐惧泛化的神经机制及其行为反应, 发现基底外侧核(basolateral complex of the amygdala, BLA)处神经元的特异变化减弱了泛化阶段的辨别学习, 这种杏仁核部位的轻微改变影响了前额叶皮层以及海马, 并破坏了泛化的平衡, 从而导致了更广的泛化.另外, 内侧前额叶(mPFC)-BLA通路的增强可以促进恐惧消退, 这种自上而下连接不良的传递, 也会导致更广的泛化(Jasnow et al., 2017; Likhtik & Paz, 2015).由此可见, 杏仁核在知觉恐惧泛化回路中有着重要的作用, 是其他脑区与感知觉信息的连接点, 控制着恐惧反应的表达. ...
Auditory aversive learning increases discrimination thresholds
1
2011
... 从研究方法上来说, 刺激的不同设置方式是否会影响被试的辨别力?首先, 在对GS恐惧程度进行评估时, 刺激单独呈现, 并被较长的时间间隔分开, 这就减弱了刺激间直接的对比.此外, 不同的时间间隔将会触发不同的记忆过程, 因为随着时间间隔的增长, 被试的行为反应会相应减弱(Burton & Sinclair, 2000).其次, 被试的情绪状态也会影响实验结果.在单纯的知觉辨别任务中, 没有呈现厌恶刺激时被试的情绪是相对中性的.但在恐惧泛化任务中, 恐惧的情绪状态会影响被试的决策并减少知觉的准确性, 从而影响被试对GS和CS的辨别(Stefanucci & Storbeck, 2009).由此可见, 个体在恐惧状态和正常状态下的辨别能力是存在差异的.再次, 刺激梯度通常是人为操纵, 而非基于被试的知觉, 而个体的感知阈限不同, 对刺激的辨别度也存在差异, 如Resnik, Sobel和Paz (2011)用纯音(1/2 kHz)分别作为CS+和CS-, 被试却经常把CS+周围的刺激认作CS+.然而, 只有极少数研究用差别阈限法关注被试对材料的辨别度(Holt et al., 2014).所以未来的研究应该注意区分被试对刺激的辨别力, 采用差别阈限法对实验材料进行设置以避免因混淆辨别力不佳而造成的实验结果误差. ...
Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation
1
2011
... 海马中 “模式分离(pattern separation)”和“模式完成(pattern completion)”负责对刺激的属性特征与已存储的信息进行匹配(Gluck & Myers, 1993).“模式分离”由海马的齿状回执行(dentate gyrus, DG), 是一种把相似信息区分开的神经活动; 而“模式完成”主要发生在海马CA3区, 它描述了相反的加工过程, 如果刺激的区别较小, “模式完成”则会忽略刺激间的区别(Drew & Huckleberry, 2017; Gilbert, Kesner, & Lee, 2001; Treves & Rolls, 1992).在知觉泛化过程中, 当GS和CS相似性较高时, 海马启动“模式完成”使CS的神经表征激活, 从而进一步引发被试的恐惧反应; 而当GS和CS相似性较低时, 海马则启动“模式分离”激活腹内测前额叶从而降低杏仁核的活动, 达到抑制恐惧反应的效果(Lissek, 2012; Lissek, Bradford et al., 2014).例如, 研究发现在啮齿动物的情景辨别恐惧泛化任务中, 海马CA3区的神经抑制会促进啮齿动物在恐惧和中性情景下的“模式完成”过程, 从而增加恐惧的泛化; 反过来, 齿状回处的神经兴奋则会促进“模式分离”过程, 以抑制恐惧泛化(Sahay et al., 2011; Niibori et al., 2012).可见, 海马在知觉恐惧泛化的过程中负责刺激的辨别加工. ...
Convergence of autonomic and limbic connections in the insular cortex of the rat
1
1982
... 脑岛和海马、杏仁核、水管周灰质(periaqueductal gray )共同控制自主神经系统, 它们在加工恐惧情绪时被激活(Saper, 1982).前人研究表明, 当个体面对不确定的情景及对恐惧刺激进行预期时, 脑岛的响应随之增强(Berns et al., 2006; Dunsmoor, Bandettini, & Knight, 2007).一般认为, 前脑岛是躯体感觉和内脏感觉的结合点, 与ACC、杏仁核、辅助运动区(supplementary motor area, SMA)等区域一起引发与恐惧相关的行为.Dunsmoor, Prince等(2011)首次用功能成像技术结合情绪面孔感知维度对人类恐惧泛化进行研究, 并证明了在条件性恐惧的习得和表达中, 当GS接近CS时, 脑岛、尾状核(caudate)、丘脑的激活程度普遍增加.例如Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013a)在用几何图形测试人类恐惧泛化的过程中, 发现在接近CS时, 脑岛、尾状核、ACC、SMA的响应会随着刺激相似性的增加而增强.随后, Greenberg, Carlson, Cha, Hajcak和Mujica-Parodi (2013b)用相同的范式对广泛性焦虑障碍患者进行恐惧泛化测试, 发现前脑岛(anterior insular, AI)与ACC、杏仁核、SMA连接增强.同时, Lissek, Bradford 等(2014)用圆圈刺激发现在恐惧泛化任务中, 正常被试脑岛激活增强.此后, Onat和Büchel (2015)用面孔范式发现脑岛对CS+最敏感, 颞下皮层(inferotemporal cortex, ITC)对CS+和CS-中间的模糊刺激比较敏感, 这表明前脑岛皮层编码“危险识别”过程, 颞下皮层则对“模糊不清”的信息进行编码.可见, 脑岛在知觉的恐惧泛化过程中参与恐惧的表达和刺激的辨别. ...
Activity of the nitrergic system of the medial prefrontal cortex in rats with high and low levels of generalization of a conditioned reflex fear reaction
1
2016
... 内侧前额叶皮层(medial prefrontal cortex, mPFC)是恐惧神经环路的关键脑区, 在恐惧消退过程中起着抑制恐惧的作用(Milad et al., 2007; Orsini & Maren, 2012).近来研究表明, mPFC与恐惧泛化过程也有密切的联系.Cullen, Gilman, Winiecki, Riccio和Jasnow (2015)发现, 情景泛化记忆的表达与前扣带皮层(anterior cingulate cortex, ACC)、下边缘皮层(infralimbic cortex, IL)、前边缘皮层(prelimbic cortex, PL)这些前额叶皮层脑区的活性有关, 其中ACC与海马CA1区的连接抑制将减弱恐惧的泛化.另外, 与脑岛等脑区相反的是, vmPFC在接近CS-时激活更强(Greenberg et al., 2013a, 2013b; Lissek, Brandford et al., 2014), 在安全线索条件下GAD患者的vmPFC激活较正常人弱(Greenberg et al., 2013b).此外, vmPFC皮质厚度和恐惧泛化梯度有关, vmPFC皮质的减少可能是导致GAD患者过度泛化的原因之一(Cha et al, 2014).Xu等(2012)观察到mPFC处突触传递的毁损将导致动物情景恐惧记忆的过度泛化.他们认为mPFC、海马以及连接核(nucleus reuniens, NR) (丘脑中线核群的一部分)共同控制着恐惧记忆的泛化.具体而言, 由于mPFC是参与动机及情绪状态的关键脑区, 因此mPFC-NR路径将记忆的动机以及情绪效价信息传递至海马进行特征的重叠编码, 海马又将编码后的信息返回至mPFC进而对泛化记忆进行调节(Xu & Südhof, 2013).另有研究表明, 老鼠对声音习得的恐惧反应和mPFC处胆碱能系统(nitrergic system) (神经元轴突分泌的一类神经递质)的激活水平能够预测泛化的程度, 表现为胆碱能系统激活程度越高, 泛化程度越高(Saul’skaya & Sudorgina, 2016).此外, Likhtik, Stujenske, Topiwala, Harris和Gordon (2014)认为内侧前额叶(mPFC)以及基底外侧核(BLA)连接的增强预示着内侧前额叶可能会影响基底外侧核的活动, 并通过抑制杏仁核的活动来减弱恐惧反应.相反的, mPFC-BLA连接的减弱则会导致对杏仁核的抑制失败, 从而增强恐惧反应.可见, mPFC在知觉恐惧泛化过程中参与调节和抑制恐惧. ...
Developmental aspects of fear: Comparing the acquisition and generalization of conditioned fear in children and adults
1
2016
... 第三类研究则用两张中性的面孔作为CS+和CS-, 将恐惧表情的面孔与尖叫声同时作为US, 来探索儿童和青少年群体的恐惧泛化规律.Lau等(2008)最先采用面孔范式对12~13岁患焦虑症的青少年进行恐惧泛化研究, 结果发现患焦虑症的青少年的泛化程度较正常组更为严重.随后, Glenn等 (2012)用该范式对8~10岁、11~13岁两个年龄阶段的儿童进行恐惧泛化的测试, 结果发现, 年长被试能够较准确地习得恐惧且泛化梯度较有规律; 而年幼被试则在泛化阶段表现出对CS-而非GS更强的恐惧反应, 这说明年长儿童对危险刺激更具有分辨力.此外, Schiele等(2016)通过对比儿童和成人的恐惧泛化, 发现儿童较成人在唤醒度和皮肤电上都显示出了更多的泛化.Schieler认为泛化梯度具有由浅入深的发展性, 很可能与大脑结构的成熟度有关, 并且大脑结构的成熟度可以调节对危险和安全线索的辨别. ...
retrograde amnesia and memory consolidation: A neurobiological perspective
1
1995
... 根据系统整合理论(systems theory), 海马通过对比新输入的感觉信息和已经存储的信息, 将其进行整合之后传输至大脑皮层, 再由大脑皮层提取记忆中的共同特征, 从而引导机体做出反应(Squire & Alvarez, 1995), 泛化的记忆正是源于记忆特征的重叠(Xu & Südhof, 2013).研究表明, 在情景恐惧泛化的范式下, 老鼠能否成功辨别危险情景和新异情景与海马活动水平密切相关, 海马细胞活性的降低将直接导致更多的泛化, 因此海马对检索详细的情景记忆信息起着至关重要的作用(Wiltgen et al., 2010). ...
Don't look down: Emotional arousal elevates height perception
1
2009
... 从研究方法上来说, 刺激的不同设置方式是否会影响被试的辨别力?首先, 在对GS恐惧程度进行评估时, 刺激单独呈现, 并被较长的时间间隔分开, 这就减弱了刺激间直接的对比.此外, 不同的时间间隔将会触发不同的记忆过程, 因为随着时间间隔的增长, 被试的行为反应会相应减弱(Burton & Sinclair, 2000).其次, 被试的情绪状态也会影响实验结果.在单纯的知觉辨别任务中, 没有呈现厌恶刺激时被试的情绪是相对中性的.但在恐惧泛化任务中, 恐惧的情绪状态会影响被试的决策并减少知觉的准确性, 从而影响被试对GS和CS的辨别(Stefanucci & Storbeck, 2009).由此可见, 个体在恐惧状态和正常状态下的辨别能力是存在差异的.再次, 刺激梯度通常是人为操纵, 而非基于被试的知觉, 而个体的感知阈限不同, 对刺激的辨别度也存在差异, 如Resnik, Sobel和Paz (2011)用纯音(1/2 kHz)分别作为CS+和CS-, 被试却经常把CS+周围的刺激认作CS+.然而, 只有极少数研究用差别阈限法关注被试对材料的辨别度(Holt et al., 2014).所以未来的研究应该注意区分被试对刺激的辨别力, 采用差别阈限法对实验材料进行设置以避免因混淆辨别力不佳而造成的实验结果误差. ...
The biology of fear- and anxiety-related behaviors
1
2002
... 恐惧是一种对环境的适应性机制, 当危险发生时, 恐惧使得机体启动防御机制并对危险刺激进行反应(Dymond, Dunsmoor, Vervliet, Roche, & Hermans, 2015; Hoppenbrouwers, Bulten, & Brazil, 2016; LeDoux, 2000).人类的恐惧反应多是后天习得, 例如, 在华生的条件反射实验中, 小阿尔伯特原本并不惧怕白鼠, 然而当白鼠与巨大的金属响声(危险刺激)同时出现后, 小阿尔伯特便对白鼠产生了恐惧反应, 此后当他再见到小白兔、白色绒毛玩具、甚至老人的白胡须等相关事物时, 他都会感到害怕(Watson & Rayner, 1920).这种不仅对特定的刺激(白鼠)产生恐惧, 而且对与特定刺激相关的事物同样会产生恐惧的现象即是“恐惧泛化” (Dunsmoor, Prince, Murty, Kragel, & LaBar, 2011; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014; Onat & Büchel, 2015).一般来说, 适当的恐惧泛化对个体生存具有积极意义, 因为在漫长的进化过程中, 恐惧泛化能够促使人类躲避各种潜在威胁, 有利于个体生存繁衍; 而在日常生活中, 恐惧泛化也可增强个体的警觉性, 从而有效并快速地对危险环境做出预测和应对(Steimer, 2002). ...
Perceptual discrimination in fear generalization: Mechanistic and clinical implications
1
2015
... 大量恐惧泛化的研究表明, 相同知觉维度的刺激可以根据物理上的相似性由高到低被分成几个不同的层次, 而个体的恐惧反应也随着刺激的相似性高低而呈现出不同反应大小的梯度 (Ghirlanda & Enquist, 2003; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014).在知觉恐惧泛化研究中, 由一个感觉刺激(记为CS+; 如特定的灯光、声音、形状)与恐惧刺激(US; 如电击、恐惧图片、尖叫声)建立的条件反应, 另一个区别于CS+的感觉刺激作为对照刺激从不匹配US (记为CS-; 如与CS+差别较大的灯光、声音、形状), 通常用相同知觉维度的刺激(记为GS; 如在CS+和CS-之间变化的灯光、声音、形状)进行泛化测试(Hermans, Baeyens, & Vervliet., 2013).这种基于知觉的恐惧泛化梯度可以反映辨别学习和恐惧的泛化程度(Lissek et al., 2008; Struyf, Zaman, Vervliet, & van Diest, 2015) (见图1). ...
Trial-unique, delayed nonmatching- to-location (TUNL): A novel, highly hippocampus- dependent automated touchscreen test of location memory and pattern separation
1
2010
... 情景的恐惧泛化需要对整个情景内的信息进行辨别, 这些信息通常包括空间、嗅觉、听觉和视觉信息, 而不是单一的物体特征的变化(Jasnow, Lynch, Gilman, & Riccio, 2017).首先, 在动物研究方面, 有研究者训练啮齿类动物对某一情景(CXT+)习得恐惧, 而在另外一个新异的情景(CXT-)下始终不匹配恐惧刺激, 紧接着在一个和CXT+相似的情景(G-CXT)中进行恐惧泛化测试.这些情景任务包括辨别触摸屏中物体与屏幕的距离(Mcallister, Saksida, & Bussey, 2013; Talpos, McTighe, Dias, Saksida, & Bussey, 2010)、辨别新异刺激和熟悉刺激的位置(Bekinschtein et al., 2013)、在不同形状的空间里习得恐惧(Mchugh et al., 2007)等.动物研究表明, 恐惧习得和重现刺激情景(G-CXT)之间的时间间隔越长, 恐惧泛化程度越大(Biedenkapp & Rudy, 2007).而在习得任务之前预先暴露情景则会减弱恐惧的泛化.此外, 研究者通过运用情景恐惧泛化范式, 发现啮齿动物恐惧泛化存在着性别差异, 具体表现为雌性比雄性更容易发生泛化(Keiser et al., 2017), 且泛化速度更快(Lynch, Cullen, Jasnow, & Riccio, 2013).预先暴露情景增加了雄性老鼠在习得阶段的恐惧反应, 但却减少了雌性老鼠在泛化的阶段恐惧反应(Keiser et al., 2017). ...
Computational constraints suggest the need for two distinct input systems to the hippocampal CA3 network
1
1992
... 海马中 “模式分离(pattern separation)”和“模式完成(pattern completion)”负责对刺激的属性特征与已存储的信息进行匹配(Gluck & Myers, 1993).“模式分离”由海马的齿状回执行(dentate gyrus, DG), 是一种把相似信息区分开的神经活动; 而“模式完成”主要发生在海马CA3区, 它描述了相反的加工过程, 如果刺激的区别较小, “模式完成”则会忽略刺激间的区别(Drew & Huckleberry, 2017; Gilbert, Kesner, & Lee, 2001; Treves & Rolls, 1992).在知觉泛化过程中, 当GS和CS相似性较高时, 海马启动“模式完成”使CS的神经表征激活, 从而进一步引发被试的恐惧反应; 而当GS和CS相似性较低时, 海马则启动“模式分离”激活腹内测前额叶从而降低杏仁核的活动, 达到抑制恐惧反应的效果(Lissek, 2012; Lissek, Bradford et al., 2014).例如, 研究发现在啮齿动物的情景辨别恐惧泛化任务中, 海马CA3区的神经抑制会促进啮齿动物在恐惧和中性情景下的“模式完成”过程, 从而增加恐惧的泛化; 反过来, 齿状回处的神经兴奋则会促进“模式分离”过程, 以抑制恐惧泛化(Sahay et al., 2011; Niibori et al., 2012).可见, 海马在知觉恐惧泛化的过程中负责刺激的辨别加工. ...
Fear generalization in humans: Impact of feature learning on conditioning and extinction
1
2014
... 图形形状特征变化通常结合颜色变化共同影响恐惧泛化.例如, Vervliet和Geens (2014)让被试对某一特定颜色的几何图形习得恐惧, CS-是含有CS+颜色或是形状特征的几何图形.泛化测试的结果表明, 被试对与CS-有相同特征的GS的恐惧反应较小.可见, 这项研究再次证明了CS-可以决定刺激泛化的模式.另外, Vervliet, Kindt, Vansteenwegen和Hermans (2010)发现口头指导语对恐惧泛化也有较明显的影响.他们将某一特定颜色的几何图形作为CS+, 在习得恐惧之前告知被试, 该几何图形的颜色或形状特征预示危险.结果发现被试对含有口头告知特征的GS更恐惧.还有研究者则认为该结果可能与注意和学习有关, 于是他们选择在习得之后呈现指导语, 而实验结果与前者一致(Ahmed & Lovibond 2015).可见, 认知过程在人类恐惧泛化中有着重要作用. ...
Fear generalization in humans: Impact of verbal instructions
1
2010
... 图形形状特征变化通常结合颜色变化共同影响恐惧泛化.例如, Vervliet和Geens (2014)让被试对某一特定颜色的几何图形习得恐惧, CS-是含有CS+颜色或是形状特征的几何图形.泛化测试的结果表明, 被试对与CS-有相同特征的GS的恐惧反应较小.可见, 这项研究再次证明了CS-可以决定刺激泛化的模式.另外, Vervliet, Kindt, Vansteenwegen和Hermans (2010)发现口头指导语对恐惧泛化也有较明显的影响.他们将某一特定颜色的几何图形作为CS+, 在习得恐惧之前告知被试, 该几何图形的颜色或形状特征预示危险.结果发现被试对含有口头告知特征的GS更恐惧.还有研究者则认为该结果可能与注意和学习有关, 于是他们选择在习得之后呈现指导语, 而实验结果与前者一致(Ahmed & Lovibond 2015).可见, 认知过程在人类恐惧泛化中有着重要作用. ...
Opposite effects of oxytocin and vasopressin on the emotional expression of the fear response
1
2008
... 鉴于恐惧泛化几乎是所有焦虑障碍的主要症状, 而女性又是焦虑障碍的易感人群 (McLean, Asnaani, Litz, & Hofmann, 2011), 因此分析女性激素对恐惧泛化的影响具有重要的临床意义.研究表明, 恐惧泛化的性别差异很可能是由雌二醇导致(Lynch et al., 2014), 雌二醇或通过促进海马的“模式完成”或者损害“模式分离”来扩大恐惧泛化(Lynch, Winiecki, Vanderhoof, Riccio, & Jasnow, 2016).此外, 催产素有减缓焦虑情绪的作用, 因此可能对治疗焦虑症有重要价值.近期科学家甚至发现催产素可以降低人类和动物的恐惧反应, 促进恐惧的消退(Eckstein et al., 2015; Hyun, Perry, Ganella, & Madsen, 2017; Missig, Ayers, Schulkin, & Rosen, 2010).如把催产素注射到啮齿动物的中央杏仁核(central amygdala)或者背外侧隔膜(dorso-lateral septum)可以减少在消退阶段的恐惧反应(Viviani & Stoop, 2008; Zoicas, Slattery, & Neumann, 2014).在对人类进行的研究中, 使用鼻喷催产素后, 被试mPFC的活动明显增强从而可以达到促使恐惧消退的效果. ...
Conditioned emotional re
1
1920
... 恐惧是一种对环境的适应性机制, 当危险发生时, 恐惧使得机体启动防御机制并对危险刺激进行反应(Dymond, Dunsmoor, Vervliet, Roche, & Hermans, 2015; Hoppenbrouwers, Bulten, & Brazil, 2016; LeDoux, 2000).人类的恐惧反应多是后天习得, 例如, 在华生的条件反射实验中, 小阿尔伯特原本并不惧怕白鼠, 然而当白鼠与巨大的金属响声(危险刺激)同时出现后, 小阿尔伯特便对白鼠产生了恐惧反应, 此后当他再见到小白兔、白色绒毛玩具、甚至老人的白胡须等相关事物时, 他都会感到害怕(Watson & Rayner, 1920).这种不仅对特定的刺激(白鼠)产生恐惧, 而且对与特定刺激相关的事物同样会产生恐惧的现象即是“恐惧泛化” (Dunsmoor, Prince, Murty, Kragel, & LaBar, 2011; Lissek et al., 2008, 2010; Lissek, Kaczkurkin et al., 2014; Onat & Büchel, 2015).一般来说, 适当的恐惧泛化对个体生存具有积极意义, 因为在漫长的进化过程中, 恐惧泛化能够促使人类躲避各种潜在威胁, 有利于个体生存繁衍; 而在日常生活中, 恐惧泛化也可增强个体的警觉性, 从而有效并快速地对危险环境做出预测和应对(Steimer, 2002). ...
The hippocampus plays a selective role in the retrieval of detailed contextual memories
1
2010
... 根据系统整合理论(systems theory), 海马通过对比新输入的感觉信息和已经存储的信息, 将其进行整合之后传输至大脑皮层, 再由大脑皮层提取记忆中的共同特征, 从而引导机体做出反应(Squire & Alvarez, 1995), 泛化的记忆正是源于记忆特征的重叠(Xu & Südhof, 2013).研究表明, 在情景恐惧泛化的范式下, 老鼠能否成功辨别危险情景和新异情景与海马活动水平密切相关, 海马细胞活性的降低将直接导致更多的泛化, 因此海马对检索详细的情景记忆信息起着至关重要的作用(Wiltgen et al., 2010). ...
Intersection of disgust and fear: Normative and pathological views
1
2000
... 在实验材料方面, 以往实验通常采用 “厌恶刺激”作为US和CS来进行匹配, 如“高度恼人但不疼”的电击、恐惧的面孔、尖叫声等.但“厌恶刺激”所能引发的仅仅是单纯的恐惧情绪吗?事实上, 厌恶和恐惧本是两种不同的情绪, 二者在面部表情、行为反应、生理反应以及主观体验上均存在差异(Klucken et al., 2012; Woody & Teachman, 2000).因此, 我们可以质疑以往的的研究是否仅能引发的是单纯的恐惧情绪.总的来说, 未来的研究应尽量设法采用仅能真正激发单纯恐惧情绪的刺激.既关注恐惧刺激材料的准确性, 也可尝试多样化的恐惧刺激材料(如恐惧的图片、场景、声音、激光等). ...
Distinct neuronal coding schemes in memory revealed by selective erasure of fast synchronous synaptic transmission
1
2012
... 内侧前额叶皮层(medial prefrontal cortex, mPFC)是恐惧神经环路的关键脑区, 在恐惧消退过程中起着抑制恐惧的作用(Milad et al., 2007; Orsini & Maren, 2012).近来研究表明, mPFC与恐惧泛化过程也有密切的联系.Cullen, Gilman, Winiecki, Riccio和Jasnow (2015)发现, 情景泛化记忆的表达与前扣带皮层(anterior cingulate cortex, ACC)、下边缘皮层(infralimbic cortex, IL)、前边缘皮层(prelimbic cortex, PL)这些前额叶皮层脑区的活性有关, 其中ACC与海马CA1区的连接抑制将减弱恐惧的泛化.另外, 与脑岛等脑区相反的是, vmPFC在接近CS-时激活更强(Greenberg et al., 2013a, 2013b; Lissek, Brandford et al., 2014), 在安全线索条件下GAD患者的vmPFC激活较正常人弱(Greenberg et al., 2013b).此外, vmPFC皮质厚度和恐惧泛化梯度有关, vmPFC皮质的减少可能是导致GAD患者过度泛化的原因之一(Cha et al, 2014).Xu等(2012)观察到mPFC处突触传递的毁损将导致动物情景恐惧记忆的过度泛化.他们认为mPFC、海马以及连接核(nucleus reuniens, NR) (丘脑中线核群的一部分)共同控制着恐惧记忆的泛化.具体而言, 由于mPFC是参与动机及情绪状态的关键脑区, 因此mPFC-NR路径将记忆的动机以及情绪效价信息传递至海马进行特征的重叠编码, 海马又将编码后的信息返回至mPFC进而对泛化记忆进行调节(Xu & Südhof, 2013).另有研究表明, 老鼠对声音习得的恐惧反应和mPFC处胆碱能系统(nitrergic system) (神经元轴突分泌的一类神经递质)的激活水平能够预测泛化的程度, 表现为胆碱能系统激活程度越高, 泛化程度越高(Saul’skaya & Sudorgina, 2016).此外, Likhtik, Stujenske, Topiwala, Harris和Gordon (2014)认为内侧前额叶(mPFC)以及基底外侧核(BLA)连接的增强预示着内侧前额叶可能会影响基底外侧核的活动, 并通过抑制杏仁核的活动来减弱恐惧反应.相反的, mPFC-BLA连接的减弱则会导致对杏仁核的抑制失败, 从而增强恐惧反应.可见, mPFC在知觉恐惧泛化过程中参与调节和抑制恐惧. ...
A neural circuit for memory specificity and generalization
2
2013
... 根据系统整合理论(systems theory), 海马通过对比新输入的感觉信息和已经存储的信息, 将其进行整合之后传输至大脑皮层, 再由大脑皮层提取记忆中的共同特征, 从而引导机体做出反应(Squire & Alvarez, 1995), 泛化的记忆正是源于记忆特征的重叠(Xu & Südhof, 2013).研究表明, 在情景恐惧泛化的范式下, 老鼠能否成功辨别危险情景和新异情景与海马活动水平密切相关, 海马细胞活性的降低将直接导致更多的泛化, 因此海马对检索详细的情景记忆信息起着至关重要的作用(Wiltgen et al., 2010). ...
... 内侧前额叶皮层(medial prefrontal cortex, mPFC)是恐惧神经环路的关键脑区, 在恐惧消退过程中起着抑制恐惧的作用(Milad et al., 2007; Orsini & Maren, 2012).近来研究表明, mPFC与恐惧泛化过程也有密切的联系.Cullen, Gilman, Winiecki, Riccio和Jasnow (2015)发现, 情景泛化记忆的表达与前扣带皮层(anterior cingulate cortex, ACC)、下边缘皮层(infralimbic cortex, IL)、前边缘皮层(prelimbic cortex, PL)这些前额叶皮层脑区的活性有关, 其中ACC与海马CA1区的连接抑制将减弱恐惧的泛化.另外, 与脑岛等脑区相反的是, vmPFC在接近CS-时激活更强(Greenberg et al., 2013a, 2013b; Lissek, Brandford et al., 2014), 在安全线索条件下GAD患者的vmPFC激活较正常人弱(Greenberg et al., 2013b).此外, vmPFC皮质厚度和恐惧泛化梯度有关, vmPFC皮质的减少可能是导致GAD患者过度泛化的原因之一(Cha et al, 2014).Xu等(2012)观察到mPFC处突触传递的毁损将导致动物情景恐惧记忆的过度泛化.他们认为mPFC、海马以及连接核(nucleus reuniens, NR) (丘脑中线核群的一部分)共同控制着恐惧记忆的泛化.具体而言, 由于mPFC是参与动机及情绪状态的关键脑区, 因此mPFC-NR路径将记忆的动机以及情绪效价信息传递至海马进行特征的重叠编码, 海马又将编码后的信息返回至mPFC进而对泛化记忆进行调节(Xu & Südhof, 2013).另有研究表明, 老鼠对声音习得的恐惧反应和mPFC处胆碱能系统(nitrergic system) (神经元轴突分泌的一类神经递质)的激活水平能够预测泛化的程度, 表现为胆碱能系统激活程度越高, 泛化程度越高(Saul’skaya & Sudorgina, 2016).此外, Likhtik, Stujenske, Topiwala, Harris和Gordon (2014)认为内侧前额叶(mPFC)以及基底外侧核(BLA)连接的增强预示着内侧前额叶可能会影响基底外侧核的活动, 并通过抑制杏仁核的活动来减弱恐惧反应.相反的, mPFC-BLA连接的减弱则会导致对杏仁核的抑制失败, 从而增强恐惧反应.可见, mPFC在知觉恐惧泛化过程中参与调节和抑制恐惧. ...
Brain oxytocin in social fear conditioning and its extinction: Involvement of the lateral septum
1
2014
... 鉴于恐惧泛化几乎是所有焦虑障碍的主要症状, 而女性又是焦虑障碍的易感人群 (McLean, Asnaani, Litz, & Hofmann, 2011), 因此分析女性激素对恐惧泛化的影响具有重要的临床意义.研究表明, 恐惧泛化的性别差异很可能是由雌二醇导致(Lynch et al., 2014), 雌二醇或通过促进海马的“模式完成”或者损害“模式分离”来扩大恐惧泛化(Lynch, Winiecki, Vanderhoof, Riccio, & Jasnow, 2016).此外, 催产素有减缓焦虑情绪的作用, 因此可能对治疗焦虑症有重要价值.近期科学家甚至发现催产素可以降低人类和动物的恐惧反应, 促进恐惧的消退(Eckstein et al., 2015; Hyun, Perry, Ganella, & Madsen, 2017; Missig, Ayers, Schulkin, & Rosen, 2010).如把催产素注射到啮齿动物的中央杏仁核(central amygdala)或者背外侧隔膜(dorso-lateral septum)可以减少在消退阶段的恐惧反应(Viviani & Stoop, 2008; Zoicas, Slattery, & Neumann, 2014).在对人类进行的研究中, 使用鼻喷催产素后, 被试mPFC的活动明显增强从而可以达到促使恐惧消退的效果. ...


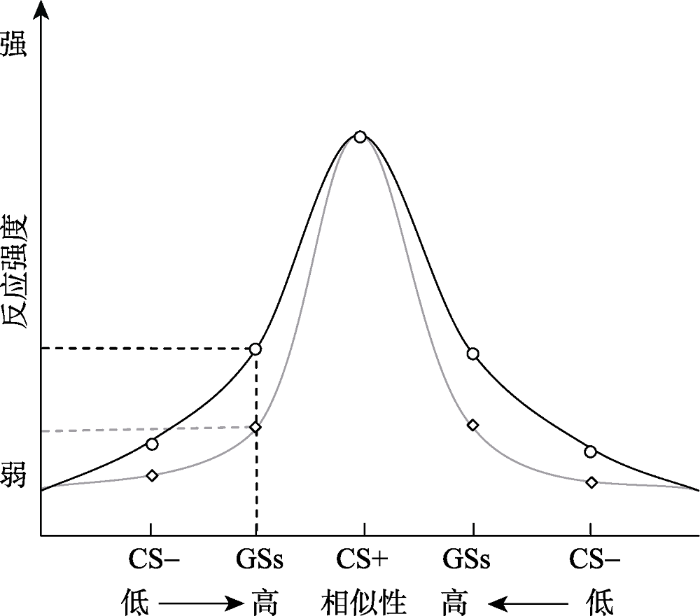
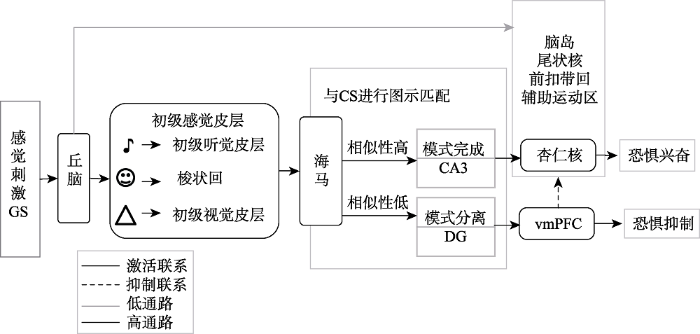

{kind=link}
{kind=link}
{kind=link}
{kind=link}