

1 引言
数学焦虑是对数学加工的不良情绪反应(Ashcraft, 2002)。数学焦虑的发生率高, 影响范围广。根据经济合作与发展组织(Organisation for Economic Co-operation and Development, OECD)的国际学生评估项目PISA 2012的统计结果, 在参与调查的65个国家和地区中, 33%的15岁青少年报告在面临数学问题时感到无助(OECD, 2013)。
高数学焦虑个体在一般性的思维和认知任务中表现正常, 但数学认知能力显著下降, 包括计算、数学问题解决等复杂数学加工(如Ashcraft & Krause, 2007), 还包括数量加工等基础数学加工(Lindskog, Winman, & Poom, 2017; Maloney, Risko, Ansari, & Fugelsang, 2010; Maloney, Ansari, & Fugelsang, 2011)。例如, 一些研究发现高数学焦虑个体在符号表达的数量加工过程中不同于低数学焦虑个体(如Dietrich, Huber, Moeller, & Klein, 2015; Maloney et al., 2010, 2011; Núñez-Peña & Suárez- Pellicioni, 2014)。Dietrich等(2015)数字大小比较任务(如比较1和2哪个数更大)中, 发现高数学焦虑组在行为上表现出更强的数字距离效应, 即高低数学焦虑个体上存在难度与组别的交互作用:对于数字距离较大的条件(如1和4)两组的行为表现差异较小, 对于数字距离较小的条件(如3和4)两组行为表现差异较大。Núñez-Peña和Suárez-Pellicioni (2014)采用数字大小比较任务(如比较1和2哪个数更大), 在行为指标和EEG波幅(200~250 ms, 中线电极Fz, Cz, Pz)都发现高数学焦虑组表现出更大的数字距离效应和数字大小效应。数字大小效应表现为, 数量较大的数字的比较(如9和10)相对于数量较小的数字的比较(如1和2)难度更大, 在高低数学焦虑个体上存在难度与组别的交互作用, 数量较小的数字的比较的组间差异低于数量较大的数字的组间差异。更强的数字距离效应与数量大小效应均表明高数学焦虑个体的数量加工能力下降。
为什么数学焦虑个体的数学能力下降?“干扰理论” (Disruption Account)认为高数学焦虑是由于短暂的认知资源的过度占用(主要是工作记忆的过度占用), 导致数学能力下降(Ashcraft & Kirk, 2001)。高数学焦虑个体在完成数学任务的同时还需要额外地处理焦虑带来的负性情绪, 而数学任务常常依赖于工作记忆资源, 情绪上的负担会影响工作记忆进而影响其完成数学任务的效率。另一种“能力减退理论” (Reduced Competency Account)认为高数学焦虑是直接由其低数学能力导致的(Carey, Hill, Devine, & Szücs, 2016; Maloney et al., 2010, 2011)。
近似数量加工能力是基本数量感知(Number Sense)能力的核心内容之一, 是复杂数学加工如计算、推理等高级数学能力的基础, 对数学学业成就也具有显著的预测作用(如Halberda, Mazzocco, & Feigenson, 2008)。作为一种非符号数量表征, 从认知发展的角度, 近似数量加工能力的发展先于语言和数字符号。动物和人类婴儿都具备对近似数量进行估计和比较的能力(Brannon, Jordan, & Jones, 2010; Izard, Sann, Spelke, & Streri, 2009; Xu, Spelke, & Goddard, 2005)。因此近似数量加工过程具有不受工作记忆的影响, 且较少受教育文化因素干扰等特点(Sullivan, Frank, & Barner, 2016)。探讨数学焦虑个体的基础数学加工(如近似数量加工)的特点有助于理解其在高级数学加工(如计算、数学问题解决)上成绩下降的原因。
那么, 数学焦虑个体的近似数量加工能力是否下降?当前行为研究的结果揭示高数学焦虑个体可能存在近似数量加工的问题。Lindskog等(2017)发现近似数量加工能力受数学焦虑水平的影响:对79名健康大学生进行行为测试, 采用相关分析与偏相关分析, 发现在控制个体视觉扫描速度及一般智力的影响前后, 个体数学焦虑水平与近似数量加工水平均呈显著负相关。但目前尚无探讨数学焦虑个体近似数量加工的神经机制的相关研究, 本研究拟为此提供来自脑电(EEG)的证据。
P2p成分被认为是近似数量加工的一个重要指标(Fornaciai, Brannon, Woldorff, & Park, 2017; Hyde, & Spelke, 2009, 2012; Libertus, Woldorff, & Brannon 2007; Park, DeWind, Woldorff, & Brannon, 2015), 是刺激发生后200 ms左右的正走向波。Libertu等(2007)首先发现P2p波幅可以被非符号的数字距离大小所调节。他们让被试比较屏幕上呈现的点阵(大小在1~4和6~9之间)的数量与5的大小。屏幕上只呈现点阵, 发现与5距离更大的点阵相比于距离小的点阵在P2p成分上诱发的波幅更大。Hyde等(2009)采用适应范式(即被动观看任务):首先序列呈现4张相同数量的点阵图, 然后呈现一张相同或不同的数量的点阵图, 并控制该图片与前4张图片数量的比例, 被试只需认真观看屏幕上图片。该研究发现大脑枕区电极上的P2p成分的波幅会受到数量比例的调节。Hyde等(2012)重复了这一发现, 并通过溯源的方法将P2p成分定位在右侧顶内沟。Park等(2015)更加系统地控制了点阵的其他视觉属性(如单个点的面积、点的稀疏度), 采用与Hyde等(2009)相同的适应范式, 基于回归分析的方法发现顶枕区电极上的P2p成分波幅大小会受到数量变化的调节, 同时该成分的波幅不受到其他视觉属性的调节。上述研究提示P2p波幅与近似数量加工的精确度有关, 精确度越高P2p波幅越大。如果高数学焦虑个体存在近似数量加工表征的问题, 则可预期高低数学焦虑个体的P2p波幅显著高于低数学焦虑个体。
除了锁时锁相的事件相关电位(Event-related Potential, ERP)之外, 大脑的神经振荡也为控制神经元的放电时间提供一种有效的度量指标(Engel & Fries, 2010)。从方法学上讲, 时频分析可以用来评估与事件相关的神经振荡信息, 揭示与刺激呈现锁时的EEG节律能量变化, 从而推测这些EEG活动的能量变化所反映的神经元节律性的调整过程(兴奋或抑制)。时频分析是一种揭示事件相关神经振荡的常用方法, 目前已有两项探讨近似数量加工神经振荡特点的相关研究。例如, Libertus和Brannon (2009)发现7个月大的婴儿在观看点阵图片时, α频段(8~12 Hz)和θ频段(4~6 Hz)的神经振荡强度在新异点阵和熟悉点阵间变化时会产生变化, 且α频段神经振荡强度可受新异点阵和熟悉点阵间数量大小的比例调节; Park (2018)采用稳态视觉诱发电位技术(Steady-State Visual Evoked Potential, SSVEP), 以8 Hz的频率呈现点阵, 并以1 Hz的频率改变点的数量、大小、稀疏程度等变量, 结果发现在1 Hz频率(δ频段, 1~3 Hz)下, 3~10岁的儿童被试和成年被试观看点阵所诱发的神经振荡强度会被点的数量变化所调节, 而不被点的其他视觉属性(点的面积、稀疏度等)的变化所调节, 该研究说明δ频段的神经振荡频率可被近似数量加工中的数量比例所调节, 而不受其他视觉属性的影响, 而Libertus等(2009)中α频段的显著作用可能是由点阵的视觉属性而不是数量属性导致的。我们假设, 如果特定频段EEG振荡的能量能被数量所调节, 且高数学焦虑个体存在近似数量加工能力下降, 则可能观察到高低数学焦虑个体在该频段的振荡能量上存在差异。
综上, 来自脑电研究的证据表明P2p成分与近似数量加工密切相关, 且低频段的神经振荡强度也可被近似数量加工所调节(Fornaciai et al., 2017; Hyde et al., 2009, 2012; Libertus et al., 2007, 2009; Park et al., 2015; Park, 2018)。本研究通过认知行为测验, 匹配高低数学焦虑两组被试的基础视觉加工速度、视觉注意能力、智力等一般认知能力及一般性焦虑水平, 并分别记录了高低数学焦虑个体在完成近似数量加工任务时的脑电信号, 之后对其进行时域和频域分析。我们假设:如果高数学焦虑个体存在近似数量加工能力下降, 则可预期高低数学焦虑个体P2p成分的平均波幅存在显著差异, 并且高低数学焦虑个体在低频段的神经振荡上也存在显著差异。
2 方法
2.1 被试
通过G*power 3.1计算(Faul, Erdfelder, Lang, & Buchner, 2007), 探测重复测量方差分析中的被试间、被试内及其交互作用, 使其统计检验力达到0.7及中等效应量(r = 0.25)需要的总样本数为64, 每组32人。我们招募了62名大学生被试参与了EEG实验。被试是来自35个不同专业的本科生。数学焦虑分组通过中文版数学焦虑量表(Mathematics Anxiety Rating Scale, MARS)得分来确定。在所有参与量表填写的被试中(N = 341), 选取百分等级为80%以上的被试为高数学焦虑组, 选取百分等级在20%以下的被试作为低数学焦虑组。同时匹配两组被试在一般性焦虑水平、视觉加工速度、视觉注意广度及智力测验上的平均成绩, 保证3个测验在两组被试间平均成绩无显著差异。由此得到高数学焦虑组被试31名, 低数学焦虑组被试31名。其中1名被试因故未完成实验, 1名被试因EEG数据噪音过太大而删除, 留下31名高数学焦虑被试和29名低数学焦虑被试。
被试人口学信息及认知测验成绩见表1。高低数学焦虑得分差异显著, t(58) = 29.07, p < 0.001, Cohen's d = 1.39; 一般性焦虑得分差异不显著, t(58) = 0.20, p = 0.84。实验开始前被试填写知情同意书。
表1 高低数学焦虑组的年龄、性别、数学焦虑、一般焦虑、视觉加工速度、视觉注意广度及的平均得分及标准差
| 组别 | 年龄(岁) | 性别(男/女) | 数学焦虑 | 一般焦虑 | 视觉加工速度 | 视觉注意广度 | 智力 |
|---|---|---|---|---|---|---|---|
| 高数学焦虑 (High math anxious, HMA) | 20.68 (1.62) | 15/16 | 94.35 (7.37) | 44.70 (5.35) | 74.06 (17.24) | 58.71 (18.78) | 5.03 (1.38) |
| 低数学焦虑 (Low math anxious, LMA) | 20.69 (1.85) | 17/12 | 41.86 (6.56) | 44.45 (4.84) | 75.86 (16.19) | 63.96 (18.91) | 5.14 (0.88) |
| t | -- | -- | 29.07 | 0.2 | -0.42 | -1.08 | -0.35 |
| p | -- | -- | <.001 | 0.84 | 0.68 | 0.28 | 0.72 |
注:括号中为标准差
2.2 被试分组及筛选的材料、任务及程序
确定入组被试的流程是:首先有高低数学焦虑各40名在在线心理学实验平台上(
图1

2.2.1 数学焦虑量表及一般性焦虑量表
采用中文版数学焦虑量表(MARS)评估被试的主观数学焦虑水平, 共25题。该量表翻译自英文版的简略数学焦虑量表(Alexander & Martray, 1989), 由两名心理学专业研究生分别对MARS进行单独翻译, 对比两份译稿形成统一的中文版的MARS。呈现可能导致数学焦虑的情境(例如, 想象即将到来的数学考试)来衡量数学焦虑的强度。要求被试在5分里克特量表上从1分(无焦虑)到5分(高度焦虑)做出反应, 以表明他们对情况的感受。用测验总分表示数学焦虑的水平。MARS的分半信度为0.83, 克隆巴赫系数(Cronbach α)为0.79。被试的一般性焦虑水平通过特质焦虑量表(STAI-T; Spielberger, Gorsuch, Lushene, Vagg, & Jacobs, 1983)进行评估, 特质焦虑量表共20题, 分半信度为0.88, 克隆巴赫系数(Cronbach α)为0.85。
2.2.2 非言语矩阵推理
非言语矩阵推理测试是基于Raven的标准渐进矩阵(Raven, Raven, & Court, 1998)设计的, 用于评估一般智力。在这项测试中, 要求被试根据图片的固有规则识别图像中缺失的部分。有6到8个候选答案。被试使用鼠标选择丢失的部分。将被试的标准9分记为最终成绩。本测验分半信度为0.81, 克隆巴赫系数(Cronbach α)为0.56。
2.2.3 快速视知觉图形匹配
本测验改编自Ekstrom, French和Harman (1976)认知测验手册中的图片辨别测验(Ekstrom et al., 1976), 旨在考查被试的快速视觉加工能力。该测验为非限时测验, 由120个不规则图形组成120道题目, 分3段完成, 每部分包括40道题目, 中间给被试足够时间休息, 要求被试完成全部题目。每道题包括左边一个目标图片和右边3个选择图片, 同时出现在屏幕中央, 持续400 ms。间隔1000 ms黑屏后出现下一道题目。要求被试又快又准地完成所有题目。将被试累计回答正确的个数记为最终的成绩。本测验分半信度为0.92, 克隆巴赫系数(Cronbach α)为0.93。
2.2.4 视觉搜索
视觉搜索任务D2改编自Bates等人视觉注意测验(Bates & Lemay, 2004)。要求被试仔细扫描每一行字符, 找到字符中有带有两条短线的d, 这两条短线可以在一起, 也可以是分开的。如果图中有带两条短线的d就按P键, 没有就按Q键。本测验共240个试次, 要求被试又快又准地完成所有题目。将被试累计回答正确的个数记为最终的成绩。本测验分半信度为0.97, 克隆巴赫系数(Cronbach α)为0.89。
2.3 EEG实验材料、任务、程序
以往探讨近似数量加工过程的神经机制时通常使用主动的数量加工(例如Libertus et al., 2007)和被动的数量加工(例如Libertus et al., 2009中的适应任务)两种范式, 本研究也囊括了主动和被动数量加工两种任务。在主动数量加工任务中, 要求被试判断刺激图片中是黄点多还是蓝点多; 在被动数量加工任务中, 为了让被试保持警觉, 要求被试判断刺激图片中的唯一的正方形是黄色还是蓝色(见图2)。
图2
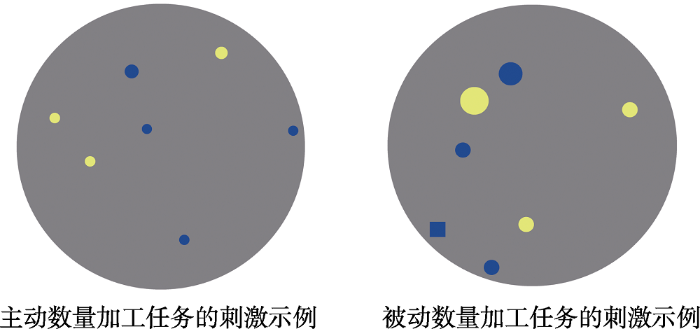
实验中, 被试坐在距离屏幕80 cm的位置平视电脑屏幕。任务呈现的程序使用E-prime 2.0 (Psychology Software Tools Inc., Sharpsburg, PA, USA)编写。在每个试次中, 首先在屏幕正中间呈现一个红色的“+”或“■”的提示800 ms, 如果呈现红色提示“+”, 被试需完成主动数量加工任务, 如果呈现红色“■”, 被试需完成被动数量加工任务。1500 ms的空屏之后呈现图片刺激, 图片刺激呈现时长为300 ms以防止被试用数数策略, 然后呈现2000 ms的空屏, 被试按“F”、“J”键进行反应, 试次间隔时间在500~2000 ms中随机。所有呈现条件在被试内平衡, 反应手在被试间平衡。实验共分为4个block, 每个block结束后有短暂休息时间。每个block中有96个试次, 每4个试次组合成一个小block, 在小block内的4个试次是同类型的任务, 即同样为主动数量加工任务或同样为被动数量加工任务, 在小block之间的试次是不同类型的任务。小block的呈现顺序是随机的, 不同的条件在小block及大block内平衡。正式实验前, 让被试进行充分的练习, 熟悉任务之后再进行正式实验。
实验材料是384张点阵图片, 主动数量加工和被动数量加工任务各192张。主动数量加工任务的实验材料主要由黄色和蓝色圆点组成, 这些点随机分布在半径固定的灰色圆型背景上。为尽量减少其他无关视觉因素对数量加工的影响, 一半的试次匹配了黄蓝颜色点的总面积, 一半试次匹配了黄蓝颜色点的平均面积(如Halberda et al., 2008)。根据韦伯定律, 数量间差异的区分主要在于比例大小, 而不是绝对差异的大小(Brannon, 2006; Hauser, Tsao, Garcia, & Spelke, 2003)。因此本研究区分了大比例数量和小比例数量两种条件, 其中大比例在2~3之间, 小比例在1.1~1.5之间, 涉及的数字范围为1~16。被动数量加工图片的条件设置与主动数量加工图片基本一致, 不同之处在于其中有一个随机点为正方形, 每张图片的视角为3°。被动数量加工任务的难度通过改变随机点的视角范围来控制。
2.4 脑电记录与分析
采用国际标准10 ~ 20系统扩展的64通道的电极帽, 用Brain Product系统记录EEG信号。滤波带通为0.01~100 Hz的频率过滤, AC采样, 采样频率为1000赫兹/导, 所有电极与头皮间的阻抗均小于10 kΩ。实验过程中以FCz作为参考电极, 离线分析时采用全脑平均参考。同时记录垂直眼电(VEOG)信号, 以监测眼球运动和眨眼。
EEG数据采用EEGLAB软件进行预处理(Delorme & Makeig, 2004)。首先对整段数据进行1~45 Hz频段的滤波, 然后分段, 分段的时间窗为刺激出现前1000 ms至刺激出现后600 ms。由于低频转换需要较长的时间周期, 1 Hz的低频信号至少需要1000 ms时长进行转换(Cohen, 2014), 参考前人的时频分析相关参数(如Zhang, Hu, Hung, Mouraux, & Iannetti, 2012)选取了现有的基线长度。从刺激开始到刺激出现600 ms长度选取主要参考前人关于P2p研究的时间窗选取参数(如Park et al., 2015)。采用EEGLAB软件中自带的“自动拒绝分段(automatic epoch rejection)”功能进行伪迹去除, 伪迹去除参数为软件的默认参数。然后用独立成分分析法(Independent component analysis, ICA)进行眼电和肌电等伪迹的去除。本研究中所涉及的被动数量加工任务是一种内隐的数量加工, 在脑电指标上应与主动数量加工同质, 因此在脑电的相关分析中应将“任务类型”变量纳入统计分析, 以考察该变量对数量加工中脑电指标的影响。总叠加平均之前各条件删除的平均trial数分别为主动观看大比例数量9 ± 6, 主动观看小比例数量9 ± 6, 被动观看大比例数量9 ± 7, 被动观看小比例数量10 ± 7。三因素重复测量方差分析结果显示, 保留的试次个数在组内条件之间无显著差异(ps > 0.05), 在组间因素间亦无显著差异, F(1,58) = 2.11, p = 0.15。
2.4.1 时域分析
根据前人文献, P2p成分是近似数量加工的一个重要指标(Fornaciai et al., 2017; Hyde et al., 2009, 2012; Libertus et al., 2007; Park et al., 2015), 该成分主要分布在头皮的后部的顶枕联合区, 位于视觉加工的背侧通路(Dorsal Stream) (Park et al., 2015)。据此, 本研究选取位于枕区电极的P2p成分进行ERP波形的统计分析。我们计算了时间窗为175~225 ms内所选枕区电极的平均波幅, 并进行2(比例类型:大比例数量和小比例数量) × 2(任务类型:主动加工和被动加工) × 2(被试类型:高数学焦虑和低数学焦虑)的重复测量方差分析。
2.4.2 时频分析
在时频分析中, 对于每一个被试, 我们采用固定250 ms汉宁窗的傅里叶变换, 将所有试次的EEG信号转换为时间-频率的信号。在每一个试次中, 用固定汉宁窗的傅里叶变换对时频空间上的每一个数据点进行复杂的时频估计, 时间为-1000 ms至600 ms (每毫秒1个采样点), 频率为1至45 Hz (每赫兹1个采样点)。然后将得到的频谱图在每一个频率上进行基线矫正(参考时间段为-900~-100 ms), 矫正
公式如下: $\text{ER}\left( \text{t},\text{f} \right)=\frac{\text{F}\left( \text{t},\text{f} \right)-\text{R}\left( \text{f} \right)}{\text{R}\left( \text{f} \right)}$, $\text{F}\left( \text{t},\text{f} \right)$表示在时间点t、频率f上信号的强度, $\text{R}\left( \text{f} \right)$表示在频率f上参考时间段的平均频率。先对EEG数据进行单试次分析, 再完成多试次的平均, 最终获得各条件下的振荡能量。
由于在时频方面近似数量加工的相关研究相对较少, 研究对象与本研究不同, 且结果不完全统一(Libertus et al., 2009; Park, 2018), 本研究采用了数据驱动的方法, 在1~45 Hz间所有频段、64个电极及刺激呈现开始至600 ms的时间窗上进行点对点的重复测量方差分析, 结合非参数置换检验法(Nonparametric Permutation Testing, Maris & Oostenveld, 2007)寻找感兴趣区(Region of Interest, ROI)。具体有如下4步:
(1)对EEG时频图上的每一个点进行2(任务类型:主动数量加工和被动数量加工) × 2(数量比例:大比例和小比例) × 2(被试类型:高数学焦虑和低数学焦虑)的重复测量方差分析, 其中任务类型、数量比例为组内因素, 被试类型为组间因素。本研究主要关注数量加工过程, 因此在考察被试类型和任务类型的主效应时, 需要首先具有数量比例效应, 即确认该感兴趣区是与数量加工相关的时频指标, 并以此为前提进行寻找有显著的被试类型差异和任务类型差异的感兴趣区。满足以下条件的时频点可进入后续分析:(1)数量比例的主效应显著的ROI; (2)数量比例主效应显著的前提下被试类型的主效应显著的ROI; (3)数量比例主效应显著的前提下任务类型的主效应显著的ROI; (4)数量比例与被试类型交互作用显著的ROI; (5)数量比例与任务类型交互作用显著的ROI; (6)以及三因素交互作用显著的ROI。上述6种ROI均满足初始p值小于0.01且包含30个连续的时间点。由此得到的每一个时频点上的F值可代表效应强度;
(2)为了解决点对点分析中多重比较矫正的问题(Maris & Oostenveld,, 2007), 从上一步分析中得到的在连续的30 ms及以上有显著效应的时频点可视为一个cluster。我们计算满足条件的每个cluster上的F值的和, 得到sum-F, 作为该cluster效应强度的指标;
(3)打乱被试类型的标签之后, 在时频分布中进行1000次置换检验。每次检验做与步骤(1)完全相同的三因素重复测量方差分析, 由此可得每个cluster在置换检验中的1000个sum-F。根据1000个sum-F的分布, 进行双尾的正态分布检验, 考察步骤(2)中得到的sum-F在打乱被试类型标签后的分布情况。
(4)为控制虚报率, 我们将通过了置换检验, 且sum-F值最大的cluster视作有效的感兴趣区(Region of interest, ROI)进入后续统计分析(Maris & Oostenveld, 2007)。
3 结果
3.1 行为结果
各条件下不同组别的平均准确率和反应时的均值和标准差如表2所示。由于被动数量加工任务是要求被试做颜色判断, 与数量加工无关, 因此不将被动数量加工的相关数据纳入行为分析。我们分别以准确率(ACC)和反应时(RT)为因变量, 进行了2(数量比例) × 2(被试类型)的两因素重复测量方差分析。
表2 各条件下不同组别的平均准确率和反应时
| 指标 | 组别 | 主动-大比例 | 主动小比例 | 被动-大比例 | 被动-小比例 |
|---|---|---|---|---|---|
| ACC | HMA | 0.91 (0.04) | 0.75 (0.05) | 0.80 (0.07) | 0.78 (0.07) |
| LMA | 0.92 (0.05) | 0.76 (0.06) | 0.78 (0.07) | 0.74 (0.07) | |
| RT | HMA | 620 (112) | 736 (132) | 619 (116) | 648 (110) |
| LMA | 622 (120) | 752 (133) | 642 (103) | 662 (101) |
注:括号中为标准差
当以准确率(ACC)为指标, (1)主效应:数量比例主效应显著(大比例数量 > 小比例数量, F(1, 58) = 772.50, p < 0.001, ηp2 = 0.93), 被试类型主效应不显著, F(1, 58) = 1.07, p = 0.31。(2)二阶交互:数量比例与被试类型的交互作用不显著, F(1, 58) = 0.77, p = 0.78 (见图3)。参考Lindskog等(2017)的分析方法, 我们将不同条件的准确率与数学焦虑得分进行了相关分析, 结果显示主动大比例(r = -0.16, p > 0.05)和主动小比例(r = -0.15, p > 0.05)条件下准确率均与数学焦虑得分呈负相关趋势。
图3
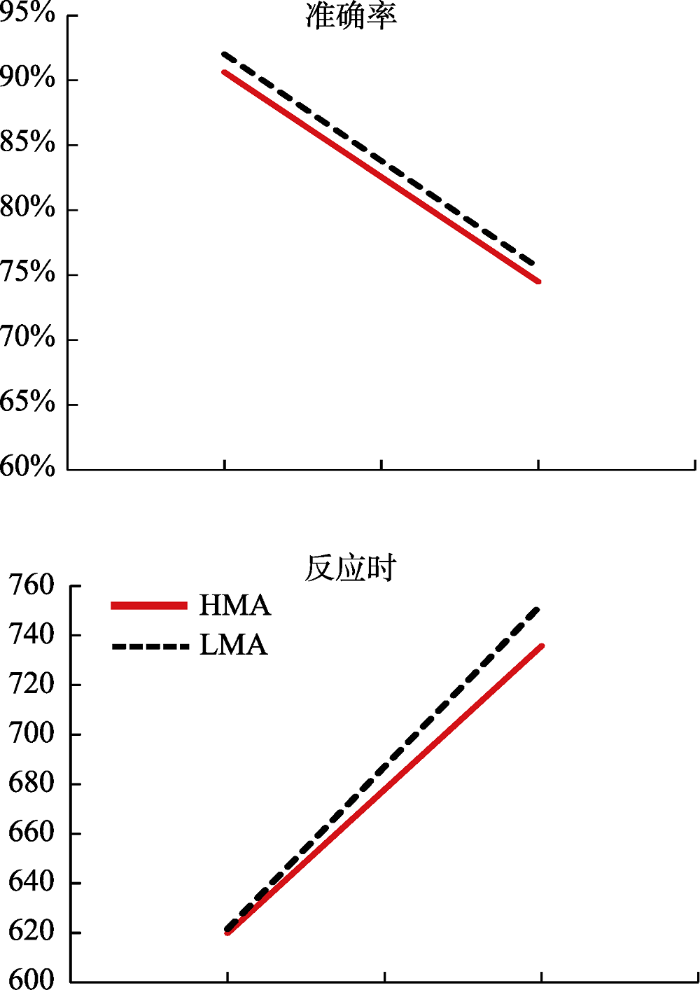
对于反应时(RT)指标, 剔除了错误反应的试次和3个标准差之外的试次之后计算每个被试在每个条件下的RT:(1)主效应:数量比例主效应显著(大比例数量 < 小比例数量, F(1, 58) = 364.78, p < 0.001, ηp2 = 0.86), 被试类型主效应不显著, F(1, 58) = 0.09, p = 0.77; (2)二阶交互:数量比例与被试类型交互作用不显著, F(1, 58) = 1.38, p = 0.25 (见图3)。
3.2 时域结果
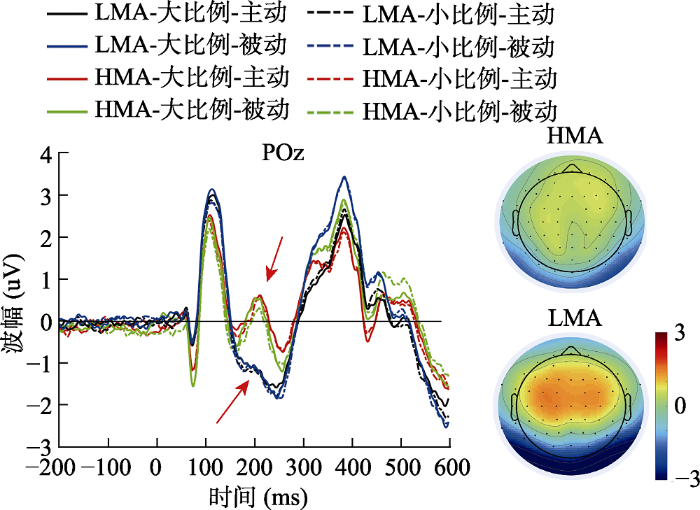
观察地形图发现, P2p成分(175~225 ms)对应的电极分布在头皮后侧(包括POz, Oz, O1, O2, PO3, PO4, PO7, PO8), 我们以这些电极点上的平均波幅为因变量, 进行2(数量比例) × 2(任务类型) × 2(被试类型)的重复测量方差分析。结果显示, (1)主效应:数量比例的主效应显著(大比例数量 > 小比例数量, F(1, 58) = 5.01, p = 0.03, ηp2 = 0.08), 被试类型主效应显著(高数学焦虑组 > 低数学焦虑组, F(1, 58) = 6.35, p = 0.01, ηp2 = 0.10), 任务类型主效应不显著, F(1, 58) = 0.59, p = 0.45; (2)三阶交互:三因素三阶交互作用不显著, F(1, 58) = 0.14, p = 0.71; (3)二阶交互, 数量比例与被试类型交互作用不显著, F(1, 58) = 0.31, p = 0.58; 任务类型与被试类型交互作用不显著, F(1, 58) = 1.01, p = 0.32; 数量比例与任务类型交互作用不显著, F(1, 58) = 1.12, p = 0.30。结果见图4。
图4
3.3 时频结果
通过点对点的非参数置换检验, 我们发现:(1)数量比例的主效应显著的1个ROI; (2)数量比例主效应显著的前提下被试类型的主效应显著的ROI不存在; (3)数量比例主效应显著的前提下任务类型的主效应显著的ROI不存在; (4)数量比例与被试类型交互作用显著的1个ROI; (5)数量比例与任务类型交互作用显著的1个ROI; (6)三因素交互作用显著的1个ROI。
数量比例主效应显著的ROI包括了AF3, AF7, AF8, F1, F3, F6, Fz, F7, C1, C2, C3, C4, C6, Cz, CP1, CP2, CP3, CP4, CPz, P1, P2, Pz和P4电极(1~6 Hz, 296~598 ms), 该ROI内神经振荡体现为事件相关同步化(Event related synchronization, ERS)。以该ROI内的平均振荡能量作为因变量, 2(数量比例) × 2(任务类型) × 2(被试类型)的重复测量方差分析显示, (1)主效应:任务类型主效应显著(主动加工 < 被动加工, F(1, 58) =7.20, p = 0.009, ηp2 = 0.11), 数量比例主效应显著(大比例数量 > 小比例数量, F(1, 58) = 23.40, p < 0.001, ηp2 = 0.29), 被试类型主效应不显著, F(1, 58) = 0.24, p = 0.62; (2)三阶交互:三因素三阶交互作用不显著, F(1, 58) = 0.36, p = 0.85; (3)二阶交互:数量比例与被试类型交互作用不显著, F(1, 58) = 0.28, p = 0.60; 数量比例与任务类型交互作用不显著, F(1, 58) = 0.09, p = 0.76; 任务类型与被试类型交互作用不显著, F(1, 58) = 0.77, p = 0.38。但该效应与眼电电极上的能量变化存在高相关, 在控制掉眼电在该时频区域的能量变化之后, 数量比例效应与任务类型的主效应均不再显著。
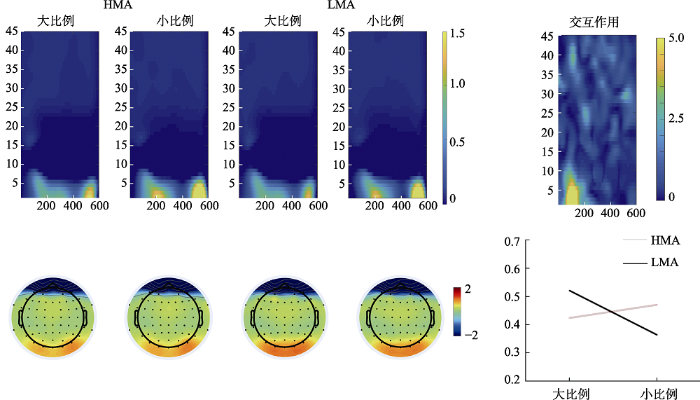
数量比例与被试类型交互作用显著的ROI包括了P5, PO7, O1和Oz电极(1~5 Hz, 83~217 ms), 该ROI内神经振荡体现为事件相关同步化(Event related synchronization, ERS) (见图5)。以该ROI内的平均振荡能量作为因变量, 2(数量比例) × 2(任务类型) × 2(被试类型)的重复测量方差分析显示, (1)主效应:任务类型主效应不显著, F(1, 58) = 0.18, p = 0.67; 数量比例主效应边缘显著(大比例数量 > 小比例数量, F(1, 58) = 3.47, p = 0.07, ηp2 = 0.06); 被试类型主效应不显著, F(1, 58) = 0, p = 0.98。(2)三阶交互:三因素三阶交互作用不显著, F(1, 58) = 0.14, p = 0.70。(3)二阶交互:数量比例与被试类型交互作用显著, F(1, 58) = 12.17, p = 0.001, ηp2 = 0.17。简单效应检验显示, 对于高数学焦虑组大小比例数量能量差异不显著, 对于低数学焦虑组, 大小比例能量差异显著(大比例数量 > 小比例数量, t(28) = 3.20, p = 0.003, Cohen's d = 0.59); 数量比例与任务类型交互作用不显著, F(1, 58) = 1.15, p = 0.70; 任务类型与被试类型交互作用不显著, F(1, 58) = 2.02, p = 0.16。
图5
数量比例与任务类型交互作用显著的ROI包括了AF3, F1, F2, F4和Fz五个电极(29~37 Hz, 48~171 ms), 该ROI内神经振荡体现为事件相关去同步化(Event related desynchronization, ERD)。以该ROI内的平均振荡能量作为因变量, 2(数量比例) × 2(任务类型) × 2(被试类型)的重复测量方差分析显示, (1)主效应:任务类型主效应不显著, F(1, 58) = 0.01, p = 0.91; 数量比例主效应不显著, F(1, 58) = 0.27, p = 0.60; 被试类型主效应不显著, F(1, 58) = 0.11,p = 0.74。(2)三阶交互:三因素三阶交互作用显著, F(1, 58) = 0.01, p = 0.91。(3)二阶交互:数量比例与被试类型交互作用不显著, F(1, 58) = 0.75, p = 0.39。任务类型与被试类型交互作用不显著, F(1, 58) = 0.23, p = 0.63。数量比例与任务类型交互作用显著, F(1, 58) = 21.37, p < 0.001, ηp2 = 0.28。简单效应显示, 在大比例数量上, 主动数量加工任务能量显著大于被动数量加工的能量, t(59) = 2.69, p = 0.009, Cohen's d = 0.35; 在小比例数量上, 主动数量加工任务能量显著小于被动数量加工的能量, t(59) = 3.13, p = 0.003, Cohen's d = 0.41。
三因素交互作用显著的ROI包括了P5和PO7两个电极(29~34 Hz, 206~285 ms), 该ROI内神经振荡体现为事件相关去同步化(Event related desynchronization, ERD)。以该ROI内的平均振荡能量作为因变量, 2(数量比例) × 2(任务类型) × 2(被试类型)的重复测量方差分析显示, (1)主效应:任务类型主效应显著(主动数量加工 > 被动数量加工, F(1, 58) = 8.63, p = 0.005, ηp2 = 0.13); 数量比例主效应不显著, F(1, 58) = 0.76, p = 0.39; 被试类型主效应不显著, F(1, 58) = 1.16, p = 0.29。(2)三阶交互:三因素三阶交互作用显著, F(1, 58) = 12.35, p = 0.001, ηp2 = 0.18。简单简单效应显示, 在高数学焦虑组的两种任务类型中, 数量比例效应均不显著(ps > 0.05); 在低数学焦虑组的主动数量加工任务中, 数量比例效应显著(大比例数量 < 小比例数量, t(28)=3.39, p = 0.002, Cohen's d = 0.63); 低数学焦虑组的被动数量加工任务中, 数量比例效应不显著(见图6)。(3)二阶交互:数量比例与被试类型交互作用不显著, F(1, 58) = 0.11, p = 0.73; 数量比例与任务类型交互作用不显著, F(1, 58) = 0.39, p = 0.54; 任务类型与被试类型交互作用显著, F(1, 58) = 5.15, p = 0.03, ηp2 = 0.08。简单效应检验显示, 高数学焦虑组在主动数量加工任务中的能量显著低于其在被动数量加工中能量, t(30)=3.65, p = 0.001, Cohen's d = 0.67; 低数学焦虑组在两种类型任务中的能量无显著差异(p > 0.05)。
图6

4 讨论
本研究考察了数学焦虑个体在近似数量加工任务中的神经电活动特点。我们发现, 在近似数量加工任务中高低数学焦虑无行为指标上的显著差异, 高数学焦虑组在枕区电极上的P2p成分平均波幅更大, 在δ频段(1~5 Hz)神经振荡上没有数量比例效应, 主动数量加工任务中β频段(29~34 Hz)神经振荡上没有数量比例效应, 而LMA在两个频段的神经振荡上均有显著的数量比例效应。这些结果为高数学焦虑个体存在近似数量加工能力下降提供了来自EEG的证据, 并为“能力减退理论”提供了数据支持。
本研究的行为结果未发现高低数学焦虑在近似数量加工中行为表现上的差异, 但是与Lindskog等人(2017)发现的相关方向是一致的, 均为负相关。本研究中主动数量加工条件的实验任务与Lindskog等(2017)的实验任务完全一致, 本研究未得到显著的负相关可能的原因有两个:第一是试次个数较少(192试次, Lindskog等(2017)中共有300试次)导致实验结果未达到稳定(Lindskog et al., 2017), 第二是任务难度较低——本研究主动条件下一半的试次为大比例数量(1 : 2和1 : 3), 而Lindskog等(2017)选用的均为小比例数量(3 : 4, 5 : 6, 7 : 8, 9 : 10)——导致在准确率上产生了一定的天花板效应。这一推测可进一步被另外两个研究的结果所证实:Dietrich等(2015)对61名高低数学焦虑个体进行行为测试, 该实验共400个试次, 设置的比例为1 : 2至9 : 10之间, 也低于Lindskog等(2017), 该研究也未发现高低数学焦虑个体在近似数量加工成绩上的显著差异。Colomé (2019)用非符号数量判断任务测试56名健康大学生, 同样未发现高低数学焦虑组在行为成绩上的组间差异, 该研究仅96个试次, 设置的比例为1 : 1.1:至1 : 2之间。结合对本研究与上述研究的实验设置我们认为, 试次的个数及任务的难度的差异可能是本研究在主动条件下两组被试在准确率上的组间差异未达统计显著的原因。但由于本实验所发现的EEG指标的理论意义需要大小数量比例对比来进行验证, 以确认该指标与近似数量加工相关, 且较多的实验试次耗时过长容易导致被试的疲劳而影响实验效果, 因此本研究为未能同时考量行为指标与EEG指标, 该推测还有待在未来研究中进一步验证。
前人研究提示高数学焦虑群体在完成数学任务会激活一些与数量加工无关的脑区激活或ERP成分, 如会激活杏仁核、脑岛等情绪、疼痛加工脑区(如Lyons & Beilock, 2011; Young, Wu, & Menon, 2012), 或在注意加工相关的P2/P3成分的波幅上存在组间差异(如Núñez-Peña & Suárez-Pellicioni, 2015; Liu, Li, Peng, Feng, & Luo, 2019)。那么本研究中P2p成分是否反映着近似数量加工过程, 能否与注意相关的P2成分相区分?首先, 本研究中P2p成分集中于枕区, 而Núñez-Peña和Suárez-Pellicioni (2015)中的P2成分的组间差异广泛地分布在额区和顶区; 其次, 本研究中枕区P2p成分的平均波幅可被数量比例所调节, 大比例数量的P2p波幅显著大于小比例数量, 这与相似范式的Libertus等人(2007)及Hyde等(2012)研究结果的方向一致。据此可推测本研究中P2p成分波幅的组间差异的确反映着近似数量的加工过程, 而不是注意加工的组间差异。Park等(2015)采用的是被动观看点阵的任务, 无需被试反应, 发现P2p波幅会随着数量的增加而增加。上述几个研究结果提示P2p波幅与近似数量加工的精确度有关, 精确度越高P2p波幅越大。P2p波幅在高数学焦虑人群中显著增强, 且在被试类型和数量比例之间无显著的交互作用, 在被试类型和任务类型之间也无显著的交互作用, 可以解释为高数学焦虑人群的近似数量加工的整体精确度有所下降。
本研究发现的显著的被试类型与数量比例的交互作用的枕区P5, PO7, O1和Oz电极的1~5 Hz上ERS出现在刺激呈现后83~217 ms, 这个频段与Park (2018)中发现的1 Hz的频段和枕区电极(Oz和PO8)的发现相吻合。Park (2018)认为在PO8上的低频神经振荡能量被数量大小所调节的效应来自于脑电的P2p成分, 而Oz上的数量效应来自于更早期的视觉加工C1成分。C1成分是被视觉刺激诱发的第一个脑电成分, 潜伏期一般是60~100 ms (Di Russo, Martínez, Sereno, Pitzalis, & Hillyard, 2002), 起源于初级视觉皮层(Fornaciai et al., 2017)。Fornaciai等(2017)发现在近似数量加工时, 在50~125 ms中诱发的脑电成分存在与C1成分类似的特点:当刺激在上下视野切换呈现时会该成分会出现极性的变化。该成分的波幅也可以显著的被数量比例所调节, 由此可推测, 在近似数量加工的早期阶段存在着一个来自初级视觉皮层的早期视觉成分也在加工数量相关的内容。而本研究中1~5 Hz的神经振荡在时间窗上与该成分有重合, 同时在低数学焦虑组中也能被数量比例调节, 由此我们推测1~5 Hz神经振荡可能来自于Fornaciai等人(2017)研究中发现的这一早期视觉成分。但由于C1成分与低频神经振荡间的关系尚未有研究证实, 且本研究的数据不能支持这一推断, 未来还需要结合严格的实验设计来证明1~5 Hz的神经振荡是否与C1成分有关。本研究在该频段上低数学焦虑组的大比例数量的能量显著大于小比例数量, 而高数学焦虑组没有显著的数量比例效应。这表明高数学焦虑个体在数量加工的早期过程对数量比例不敏感, 没有对刺激的数量信息进行充分的加工。在三阶交互作用的简单简单效应分析中, 我们也发现在枕区电极(P5和PO7)的β频段ERD (29~34 Hz, 206~285 ms), 在主动数量加工任务中, 高数学焦虑组没有表现出显著的数量比例, 而低数学焦虑组的大比例数量的能量显著低于小比例数量的能量。由于beta频段活动与自上而下的认知加工相关(见综述Engel, Fries, & Singer, 2001), β频段上的数量比例效应可能体现着自上而下的数量加工的特点, 高数学焦虑组在近似数量加工中自上而下的认知控制中可能存在问题。这是首次发现的近似数量加工的数量比例效应在高频段的EEG指标。
对于任务类型变量, 两种类型的任务都能激活P2p, 且该成分的波幅在主动数量加工和被动数量加工两种类型的任务上没有差异。以往研究分别采用了主动数量加工(Libertus et al., 2007)或被动数量加工任务(如Hyde et al., 2009, 2011, 2012; Park et al., 2015; Fornaciai et al., 2017)。这两种任务均能激活P2p成分, 但没有研究比较在不同任务中P2p成分波幅的差异。本研究同时采用了这两类任务, 发现P2p成分的波幅在两类任务中没有显著差异, 且与数量大小比例也没有显著的交互作用, 表明P2p成分是在近似数量加工中自动激活的, 可以完全独立于实验任务。由于在婴幼儿、动物实验中常采用被动数量加工范式, 但该范式缺少相应的行为反应指标, 现有结果可为被动数量加工范式的可靠性提供数据支持。存在显著的任务类型效应的时频指标体现在2个ROI上:第一个ROI是三阶交互作用显著的ROI中主动数量加工条件下神经振荡能量显著大于被动数量加工条件, 前文讨论到在该ROI内的β频段ERD可能与数量的自上而下的加工有关, 该结果可解释为主动数量加工条件中自上而下的加工更强, 因此振荡的能量更强; 第二个ROI是数量比例与任务类型交互作用显著的额区电极ROI (29~37 Hz, 48~171 ms)中, 我们发现在大比例数量上, 主动数量加工任务能量显著大于被动数量加工的能量, 在小比例数量上模式则相反, 表明支撑两种不同的任务的数量加工的EEG指标还是存在差异的, 这一差异发生在额区的beta频段可能也提示着自上而下的数量加工上的差异。
本研究的结果提示在无需工作记忆支持的近似数量加工任务中高数学焦虑的EEG指标也存在异常, 表明“干扰理论”是不准确的。以往支持“能力减退理论”的证据发现主要来自两方面:一方面关于数学焦虑发生发展的研究(Sorvo et al., 2019; Wang et al., 2014)。近期的一项纵向研究通过交叉滞后分析证明了低数学能力是因, 高数学焦虑是果(Sorvo et al., 2019)。另一方面是高数学焦虑个体基本的数量加工能力的相关研究(Lindskog et al., 2017; Maloney et al., 2010)。其基本逻辑是个体在底层的数学加工如基本数量/空间加工上的能力下降导致了高数学焦虑。现有研究发现高数学焦虑个体在符号表达的数量加工能力低于低数学焦虑个体(如Dietrich et al., 2015; Maloney et al., 2010, 2011; Núñez-Peña & Suárez-Pellicioni, 2014)。近似数量加工系统是后期习得的数量加工系统的重要组成部分, 具有不受一般认知加工及教育文化因素干扰等特点(Sullivan et al., 2016), 相对于符号表达的数量加工, 个体在近似数量系统的不精确表征更能反映被试的先天特性。Lindskog等(2017)发现数学焦虑水平与非符号的数量加工能力(即近似数量加工能力)之间存在显著的负相关。本研究在Lindskog等(2017)的基础上进一步提供了神经电生理的证据:P2p成分上显著的组间差异以及低频段(1~5 Hz)和主动数量加工中的高频段(29~34 Hz)上的交互作用均表明, 高数学焦虑个体在近似数量的加工上存在问题。高数学焦虑人群在高级的数学加工中表现较差, 可能是由于个体近似数量加工能力下降导致了高数学焦虑, 进而使得个体接受正式的数学教育之后学习时间减少、学习动机降低, 并进一步影响其数学表现。
本研究未在行为层面上发现高低数学焦虑的显著组间差异, 这是本研究的不足之处。但该结果也可进一步说明, 即使行为成绩没有显著差异, 高低数学焦虑在进行近似数量加工时仍会在与数量加工相关的神经指标上表现出异常。综上所述, 本研究为数学焦虑个体近似数量加工的能力下降的原因提供了来自EEG的证据, 支持了数学焦虑与数学能力间关系的“能力减退理论”假说。未来研究中还需增加实验难度, 结合行为结果继续验证本研究的发现; 同时还应从纵向研究的角度探讨数学焦虑、近似数量加工和数学表现三者间的相互作用, 以揭示数学焦虑、近似数量加工和数学成绩三者的因果关系。
参考文献
The development of an abbreviated version of the Mathematics Anxiety Rating Scale
Math anxiety: Personal, educational, and cognitive consequences
The relationships among working memory, math anxiety, and performance
Working memory, math performance, and math anxiety
DOI:10.3758/bf03194059
URL
PMID:17694908
[本文引用: 1]

The cognitive literature now shows how critically math performance depends on working memory, for any form of arithmetic and math that involves processes beyond simple memory retrieval. The psychometric literature is also very clear on the global consequences of mathematics anxiety. People who are highly math anxious avoid math: They avoid elective coursework in math, both in high school and college, they avoid college majors that emphasize math, and they avoid career paths that involve math. We go beyond these psychometric relationships to examine the cognitive consequences of math anxiety. We show how performance on a standardized math achievement test varies as a function of math anxiety, and that math anxiety compromises the functioning of working memory. High math anxiety works much like a dual task setting: Preoccupation with one's math fears and anxieties functions like a resource-demanding secondary task. We comment on developmental and educational factors related to math and working memory, and on factors that may contribute to the development of math anxiety.
The d2 test of attention: construct validity and extensions in scoring techniques
The representation of numerical magnitude
DOI:10.1016/j.conb.2006.03.002
URL
PMID:16546373
[本文引用: 1]
The combined efforts of many fields are advancing our understanding of how number is represented. Researchers studying numerical reasoning in adult humans, developing humans and non-human animals are using a suite of behavioral and neurobiological methods to uncover similarities and differences in how each population enumerates and compares quantities to identify the neural substrates of numerical cognition. An important picture emerging from this research is that adult humans share with non-human animals a system for representing number as language-independent mental magnitudes and that this system emerges early in development.
Behavioral signatures of numerical cognition
The chicken or the egg? The direction of the relationship between mathematics anxiety and mathematics performance
DOI:10.3389/fpsyg.2015.01987
URL
PMID:26779093
[本文引用: 1]
This review considers the two possible causal directions between mathematics anxiety (MA) and poor mathematics performance. Either poor maths performance may elicit MA (referred to as the Deficit Theory), or MA may reduce future maths performance (referred to as the Debilitating Anxiety Model). The evidence is in conflict: the Deficit Theory is supported by longitudinal studies and studies of children with mathematical learning disabilities, but the Debilitating Anxiety Model is supported by research which manipulates anxiety levels and observes a change in mathematics performance. It is suggested that this mixture of evidence might indicate a bidirectional relationship between MA and mathematics performance (the Reciprocal Theory), in which MA and mathematics performance can influence one another in a vicious cycle.
Analyzing neural time series data: Theory and practice
Representation of numerical magnitude in math-anxious individuals
EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis
DOI:10.1016/j.jneumeth.2003.10.009
URL
PMID:15102499
[本文引用: 1]
We have developed a toolbox and graphic user interface, EEGLAB, running under the crossplatform MATLAB environment (The Mathworks, Inc.) for processing collections of single-trial and/or averaged EEG data of any number of channels. Available functions include EEG data, channel and event information importing, data visualization (scrolling, scalp map and dipole model plotting, plus multi-trial ERP-image plots), preprocessing (including artifact rejection, filtering, epoch selection, and averaging), independent component analysis (ICA) and time/frequency decompositions including channel and component cross-coherence supported by bootstrap statistical methods based on data resampling. EEGLAB functions are organized into three layers. Top-layer functions allow users to interact with the data through the graphic interface without needing to use MATLAB syntax. Menu options allow users to tune the behavior of EEGLAB to available memory. Middle-layer functions allow users to customize data processing using command history and interactive 'pop' functions. Experienced MATLAB users can use EEGLAB data structures and stand-alone signal processing functions to write custom and/or batch analysis scripts. Extensive function help and tutorial information are included. A 'plug-in' facility allows easy incorporation of new EEG modules into the main menu. EEGLAB is freely available (http://www.sccn.ucsd.edu/eeglab/) under the GNU public license for noncommercial use and open source development, together with sample data, user tutorial and extensive documentation.
The influence of math anxiety on symbolic and non-symbolic magnitude processing
DOI:10.3389/fpsyg.2015.01621
URL
PMID:26579012
[本文引用: 4]
Deficits in basic numerical abilities have been investigated repeatedly as potential risk factors of math anxiety. Previous research suggested that also a deficient approximate number system (ANS), which is discussed as being the foundation for later math abilities, underlies math anxiety. However, these studies examined this hypothesis by investigating ANS acuity using a symbolic number comparison task. Recent evidence questions the view that ANS acuity can be assessed using a symbolic number comparison task. To investigate whether there is an association between math anxiety and ANS acuity, we employed both a symbolic number comparison task and a non-symbolic dot comparison task, which is currently the standard task to assess ANS acuity. We replicated previous findings regarding the association between math anxiety and the symbolic distance effect for response times. High math anxious individuals showed a larger distance effect than less math anxious individuals. However, our results revealed no association between math anxiety and ANS acuity assessed using a non-symbolic dot comparison task. Thus, our results did not provide evidence for the hypothesis that a deficient ANS underlies math anxiety. Therefore, we propose that a deficient ANS does not constitute a risk factor for the development of math anxiety. Moreover, our results suggest that previous interpretations regarding the interaction of math anxiety and the symbolic distance effect have to be updated. We suggest that impaired number comparison processes in high math anxious individuals might account for the results rather than deficient ANS representations. Finally, impaired number comparison processes might constitute a risk factor for the development of math anxiety. Implications for current models regarding the origins of math anxiety are discussed.
Cortical sources of the early components of the visual evoked potential
DOI:10.1002/hbm.10010
URL
PMID:11835601
[本文引用: 1]
This study aimed to characterize the neural generators of the early components of the visual evoked potential (VEP) to isoluminant checkerboard stimuli. Multichannel scalp recordings, retinotopic mapping and dipole modeling techniques were used to estimate the locations of the cortical sources giving rise to the early C1, P1, and N1 components. Dipole locations were matched to anatomical brain regions visualized in structural magnetic resonance imaging (MRI) and to functional MRI (fMRI) activations elicited by the same stimuli. These converging methods confirmed previous reports that the C1 component (onset latency 55 msec; peak latency 90-92 msec) was generated in the primary visual area (striate cortex; area 17). The early phase of the P1 component (onset latency 72-80 msec; peak latency 98-110 msec) was localized to sources in dorsal extrastriate cortex of the middle occipital gyrus, while the late phase of the P1 component (onset latency 110-120 msec; peak latency 136-146 msec) was localized to ventral extrastriate cortex of the fusiform gyrus. Among the N1 subcomponents, the posterior N150 could be accounted for by the same dipolar source as the early P1, while the anterior N155 was localized to a deep source in the parietal lobe. These findings clarify the anatomical origin of these VEP components, which have been studied extensively in relation to visual-perceptual processes.
Dynamic predictions: Oscillations and synchrony in top-down processing
DOI:10.1038/35094565
URL
PMID:11584308
[本文引用: 1]
Classical theories of sensory processing view the brain as a passive, stimulus-driven device. By contrast, more recent approaches emphasize the constructive nature of perception, viewing it as an active and highly selective process. Indeed, there is ample evidence that the processing of stimuli is controlled by top-down influences that strongly shape the intrinsic dynamics of thalamocortical networks and constantly create predictions about forthcoming sensory events. We discuss recent experiments indicating that such predictions might be embodied in the temporal structure of both stimulus-evoked and ongoing activity, and that synchronous oscillations are particularly important in this process. Coherence among subthreshold membrane potential fluctuations could be exploited to express selective functional relationships during states of expectancy or attention, and these dynamic patterns could allow the grouping and selection of distributed neuronal responses for further processing.
Beta-band oscillations-signalling the status quo?
DOI:10.1016/j.conb.2010.02.015
URL
PMID:20359884
[本文引用: 1]
In this review, we consider the potential functional role of beta-band oscillations, which at present is not yet well understood. We discuss evidence from recent studies on top-down mechanisms involved in cognitive processing, on the motor system and on the pathophysiology of movement disorders that suggest a unifying hypothesis: beta-band activity seems related to the maintenance of the current sensorimotor or cognitive state. We hypothesize that beta oscillations and/or coupling in the beta-band are expressed more strongly if the maintenance of the status quo is intended or predicted, than if a change is expected. Moreover, we suggest that pathological enhancement of beta-band activity is likely to result in an abnormal persistence of the status quo and a deterioration of flexible behavioural and cognitive control.
G* Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences
DOI:10.3758/BF03193146
URL
[本文引用: 1]
t, F, and χ2 test families. In addition, it includes power analyses forz tests and some exact tests. G*Power 3 provides improved effect size calculators and graphic options, supports both distribution-based and design-based input modes, and offers all types of power analyses in which users might be interested. Like its predecessors, G*Power 3 is free.]]>
Numerosity processing in early visual cortex
DOI:10.1016/j.neuroimage.2017.05.069
URL
PMID:28583882
[本文引用: 7]
While parietal cortex is thought to be critical for representing numerical magnitudes, we recently reported an event-related potential (ERP) study demonstrating selective neural sensitivity to numerosity over midline occipital sites very early in the time course, suggesting the involvement of early visual cortex in numerosity processing. However, which specific brain area underlies such early activation is not known. Here, we tested whether numerosity-sensitive neural signatures arise specifically from the initial stages of visual cortex, aiming to localize the generator of these signals by taking advantage of the distinctive folding pattern of early occipital cortices around the calcarine sulcus, which predicts an inversion of polarity of ERPs arising from these areas when stimuli are presented in the upper versus lower visual field. Dot arrays, including 8-32dots constructed systematically across various numerical and non-numerical visual attributes, were presented randomly in either the upper or lower visual hemifields. Our results show that neural responses at about 90ms post-stimulus were robustly sensitive to numerosity. Moreover, the peculiar pattern of polarity inversion of numerosity-sensitive activity at this stage suggested its generation primarily in V2 and V3. In contrast, numerosity-sensitive ERP activity at occipito-parietal channels later in the time course (210-230ms) did not show polarity inversion, indicating a subsequent processing stage in the dorsal stream. Overall, these results demonstrate that numerosity processing begins in one of the earliest stages of the cortical visual stream.
Individual differences in non-verbal number acuity correlate with maths achievement
DOI:10.1038/nature07246
URL
PMID:18776888
[本文引用: 2]
Human mathematical competence emerges from two representational systems. Competence in some domains of mathematics, such as calculus, relies on symbolic representations that are unique to humans who have undergone explicit teaching. More basic numerical intuitions are supported by an evolutionarily ancient approximate number system that is shared by adults, infants and non-human animals-these groups can all represent the approximate number of items in visual or auditory arrays without verbally counting, and use this capacity to guide everyday behaviour such as foraging. Despite the widespread nature of the approximate number system both across species and across development, it is not known whether some individuals have a more precise non-verbal 'number sense' than others. Furthermore, the extent to which this system interfaces with the formal, symbolic maths abilities that humans acquire by explicit instruction remains unknown. Here we show that there are large individual differences in the non-verbal approximation abilities of 14-year-old children, and that these individual differences in the present correlate with children's past scores on standardized maths achievement tests, extending all the way back to kindergarten. Moreover, this correlation remains significant when controlling for individual differences in other cognitive and performance factors. Our results show that individual differences in achievement in school mathematics are related to individual differences in the acuity of an evolutionarily ancient, unlearned approximate number sense. Further research will determine whether early differences in number sense acuity affect later maths learning, whether maths education enhances number sense acuity, and the extent to which tertiary factors can affect both.
Evolutionary foundations of number: spontaneous representation of numerical magnitudes by cotton-top tamarins
All numbers are not equal: an electrophysiological investigation of small and large number representations
DOI:10.1162/jocn.2009.21090
URL
PMID:18752403
[本文引用: 6]
Behavioral and brain imaging research indicates that human infants, humans adults, and many nonhuman animals represent large nonsymbolic numbers approximately, discriminating between sets with a ratio limit on accuracy. Some behavioral evidence, especially with human infants, suggests that these representations differ from representations of small numbers of objects. To investigate neural signatures of this distinction, event-related potentials were recorded as adult humans passively viewed the sequential presentation of dot arrays in an adaptation paradigm. In two studies, subjects viewed successive arrays of a single number of dots interspersed with test arrays presenting the same or a different number; numerical range (small numerical quantities 1-3 vs. large numerical quantities 8-24) and ratio difference varied across blocks as continuous variables were controlled. An early-evoked component (N1), observed over widespread posterior scalp locations, was modulated by absolute number with small, but not large, number arrays. In contrast, a later component (P2p), observed over the same scalp locations, was modulated by the ratio difference between arrays for large, but not small, numbers. Despite many years of experience with symbolic systems that apply equally to all numbers, adults spontaneously process small and large numbers differently. They appear to treat small-number arrays as individual objects to be tracked through space and time, and large-number arrays as cardinal values to be compared and manipulated.
Spatiotemporal dynamics of processing nonsymbolic number: An event‐related potential source localization study
DOI:10.1002/hbm.21352
URL
[本文引用: 6]
Coordinated studies with adults, infants, and nonhuman animals provide evidence for two distinct systems of nonverbal number representation. The parallel individuation (PI) system selects and retains information about one to three individual entities and the numerical magnitude system establishes representations of the approximate cardinal value of a group. Recent event-related potential (ERP) work has demonstrated that these systems reliably evoke functionally and temporally distinct patterns of brain response that correspond to established behavioral signatures. However, relatively little is known about the neural generators of these ERP signatures. To address this question, we targeted known ERP signatures of these systems, by contrasting processing of small versus large nonsymbolic numbers, and used a source localization algorithm (LORETA) to identify their cortical origins. Early processing of small numbers, showing the signature effects of PI on the N1 (similar to 150 ms), was localized primarily to extrastriate visual regions. In contrast, qualitatively and temporally distinct processing of large numbers, showing the signatures of approximate number representation on the mid-latency P2p (similar to 200-250 ms), was localized primarily to right intraparietal regions. In comparison, mid-latency small number processing was localized to the right temporalparietal junction and left-lateralized intraparietal regions. These results add spatial information to the emerging ERP literature documenting the process by which we represent number. Furthermore, these results substantiate recent claims that early attentional processes determine whether a collection of objects will be represented through PI or as an approximate numerical magnitude by providing evidence that downstream processing diverges to distinct cortical regions. Hum Brain Mapp 33:2189-2203, 2012. (c) 2011 Wiley Periodicals, Inc.
Spatial attention determines the nature of nonverbal number representation
DOI:10.1162/jocn.2010.21581
URL
PMID:20961170
[本文引用: 1]
Coordinated studies of adults, infants, and nonhuman animals provide evidence for two systems of nonverbal number representation: a
Newborn infants perceive abstract numbers
Behavioral and neural basis of number sense in infancy
DOI:10.1111/j.1467-8721.2009.01665.x
URL
PMID:20419075
[本文引用: 5]
Approximate number discrimination in adult human and nonhuman animals is governed by Weber's Law: The ratio between the values determines discriminability. Here, we review recent evidence from behavioral and neuroimaging studies that suggests that number sense in human infancy shares the same hallmark feature of Weber's Law and may rely on the same neural substrates as previously found in adults, children, and nonhuman animals. These findings support the notion of ontogenetic and phylogenetic continuity in number sense. New methods described here may help uncover how infants' early number sense supports the development of a mature number sense. Moreover, they may aid in understanding how children learn to map nonsymbolic number representations onto symbols for number by providing dependent measures that capture individual variability.
Electrophysiological evidence for notation independence in numerical processing
Individual differences in nonverbal number skills predict math anxiety
URL PMID:27960118 [本文引用: 12]
EEG correlates of math anxiety during arithmetic problem solving: Implication for attention deficits
DOI:10.1016/j.neulet.2019.03.047
URL
PMID:30928479
[本文引用: 1]
Anxiety about math can lead to long-term negative consequences related to academic achievement and professional success. However, it remains unclear how elevated math-anxiety modulates brain activity while solving arithmetic problems. In the current study, we recorded electrophysiological responses throughout arithmetic problem solving, both at the period of anticipating an upcoming arithmetic problem and solving an arithmetic problem. Results showed that, after controlling for mathematical performance, people with higher math anxiety tended to show stronger beta band oscillation and P300 amplitude while expecting the arithmetic problems, as well as stronger gamma band activity while solving the arithmetic problems. These results suggest that individuals highly anxious about math might use more attentional resources during the course of anticipating the upcoming arithmetic problems, and showed greater attentional bias toward arithmetic problems during solving arithmetic problems.
Mathematics anxiety: separating the math from the anxiety
DOI:10.1093/cercor/bhr289
URL
PMID:22016480
[本文引用: 1]
Anxiety about math is tied to low math grades and standardized test scores, yet not all math-anxious individuals perform equally poorly in math. We used functional magnetic resonance imaging to separate neural activity during the anticipation of doing math from activity during math performance itself. For higher (but not lower) math-anxious individuals, increased activity in frontoparietal regions when simply anticipating doing math mitigated math-specific performance deficits. This network included bilateral inferior frontal junction, a region involved in cognitive control and reappraisal of negative emotional responses. Furthermore, the relation between frontoparietal anticipatory activity and highly math-anxious individuals' math deficits was fully mediated (or accounted for) by activity in caudate, nucleus accumbens, and hippocampus during math performance. These subcortical regions are important for coordinating task demands and motivational factors during skill execution. Individual differences in how math-anxious individuals recruit cognitive control resources prior to doing math and motivational resources during math performance predict the extent of their math deficits. This work suggests that educational interventions emphasizing control of negative emotional responses to math stimuli (rather than merely additional math training) will be most effective in revealing a population of mathematically competent individuals, who might otherwise go undiscovered.
Rapid communication: the effect of mathematics anxiety on the processing of numerical magnitude
Mathematics anxiety affects counting but not subitizing during visual enumeration
DOI:10.1016/j.cognition.2009.09.013
URL
PMID:19896124
[本文引用: 7]
Individuals with mathematics anxiety have been found to differ from their non-anxious peers on measures of higher-level mathematical processes, but not simple arithmetic. The current paper examines differences between mathematics anxious and non-mathematics anxious individuals in more basic numerical processing using a visual enumeration task. This task allows for the assessment of two systems of basic number processing: subitizing and counting. Mathematics anxious individuals, relative to non-mathematics anxious individuals, showed a deficit in the counting but not in the subitizing range. Furthermore, working memory was found to mediate this group difference. These findings demonstrate that the problems associated with mathematics anxiety exist at a level more basic than would be predicted from the extant literature.
Nonparametric statistical testing of EEG and MEG data
DOI:10.1016/j.jneumeth.2007.03.024
URL
PMID:17517438
[本文引用: 3]
In this paper, we show how ElectroEncephaloGraphic (EEG) and MagnetoEncephaloGraphic (MEG) data can be analyzed statistically using nonparametric techniques. Nonparametric statistical tests offer complete freedom to the user with respect to the test statistic by means of which the experimental conditions are compared. This freedom provides a straightforward way to solve the multiple comparisons problem (MCP) and it allows to incorporate biophysically motivated constraints in the test statistic, which may drastically increase the sensitivity of the statistical test. The paper is written for two audiences: (1) empirical neuroscientists looking for the most appropriate data analysis method, and (2) methodologists interested in the theoretical concepts behind nonparametric statistical tests. For the empirical neuroscientist, a large part of the paper is written in a tutorial-like fashion, enabling neuroscientists to construct their own statistical test, maximizing the sensitivity to the expected effect. And for the methodologist, it is explained why the nonparametric test is formally correct. This means that we formulate a null hypothesis (identical probability distribution in the different experimental conditions) and show that the nonparametric test controls the false alarm rate under this null hypothesis.
Less precise representation of numerical magnitude in high math-anxious individuals: An ERP study of the size and distance effects
Processing of multi-digit additions in high math-anxious individuals: psychophysiological evidence
DOI:10.3389/fpsyg.2015.01268
URL
PMID:26347705
[本文引用: 2]
We investigated the time course of neural processing of multi-digit additions in high- (HMA) and low-math anxious (LMA) individuals. Seventeen HMA and 17 LMA individuals were presented with two-digit additions and were asked to perform a verification task. Behavioral data showed that HMA individuals were slower and more error prone than their LMA peers, and that incorrect solutions were solved more slowly and less accurately than correct ones. Moreover, HMA individuals tended to need more time and commit more errors when having to verify incorrect solutions than correct ones. ERPs time-locked to the presentation of the addends (calculation phase) and to the presentation of the proposed solution (verification phase) were also analyzed. In both phases, a P2 component of larger amplitude was found for HMA individuals than for their LMA peers. Because the P2 component is considered to be a biomarker of the mobilization of attentional resources toward emotionally negative stimuli, these results suggest that HMA individuals may have invested more attentional resources both when processing the addends (calculation phase) and when they had to report whether the proposed solution was correct or not (verification phase), as compared to their LMA peers. Moreover, in the verification phase, LMA individuals showed a larger late positive component (LPC) for incorrect solutions at parietal electrodes than their HMA counterparts. The smaller LPC shown by HMA individuals when verifying incorrect solutions suggests that these solutions may have been appeared more plausible to them than to their LMA counterparts.
PISA 2012 Results: Ready to learn: Students' engagement drive and self-beliefs
(Volume III).
A neural basis for the visual sense of number and its development: A steady-state visual evoked potential study in children and adults
DOI:10.1016/j.dcn.2017.02.011
URL
PMID:28342780
[本文引用: 5]
While recent studies in adults have demonstrated the existence of a neural mechanism for a visual sense of number, little is known about its development and whether such a mechanism exists at young ages. In the current study, I introduce a novel steady-state visual evoked potential (SSVEP) technique to objectively quantify early visual cortical sensitivity to numerical and non-numerical magnitudes of a dot array. I then examine this neural sensitivity to numerical magnitude in children between three and ten years of age and in college students. Children overall exhibit strong SSVEP sensitivity to numerical magnitude in the right occipital sites with negligible SSVEP sensitivity to non-numerical magnitudes, the pattern similar to what is observed in adults. However, a closer examination of age differences reveals that this selective neural sensitivity to numerical magnitude, which is close to absent in three-year-olds, increases steadily as a function of age, while there is virtually no neural sensitivity to other non-numerical magnitudes across these ages. These results demonstrate the emergence of a neural mechanism underlying direct perception of numerosity across early and middle childhood and provide a potential neural mechanistic explanation for the development of humans' primitive, non-verbal ability to comprehend number.
Rapid and direct encoding of numerosity in the visual stream
DOI:10.1093/cercor/bhv017
URL
PMID:25715283
[本文引用: 8]
Humans are endowed with an intuitive number sense that allows us to perceive and estimate numerosity without relying on language. It is controversial, however, as to whether there is a neural mechanism for direct perception of numerosity or whether numerosity is perceived indirectly via other perceptual properties. In this study, we used a novel regression-based analytic method, which allowed an assessment of the unique contributions of visual properties, including numerosity, to explain visual evoked potentials of participants passively viewing dot arrays. We found that the human brain is uniquely sensitive to numerosity and more sensitive to changes in numerosity than to changes in other visual properties, starting extremely early in the visual stream: 75 ms over a medial occipital site and 180 ms over bilateral occipitoparietal sites. These findings provide strong evidence for the existence of a neural mechanism for rapidly and directly extracting numerosity information in the human visual pathway.
Development of math anxiety and its longitudinal relationships with arithmetic achievement among primary school children
Intensive math training does not affect approximate number acuity: Evidence from a three-year longitudinal curriculum intervention
Who is afraid of math? Two sources of genetic variance for mathematical anxiety
DOI:10.1111/jcpp.12224
URL
PMID:24611799
[本文引用: 1]
BACKGROUND: Emerging work suggests that academic achievement may be influenced by the management of affect as well as through efficient information processing of task demands. In particular, mathematical anxiety has attracted recent attention because of its damaging psychological effects and potential associations with mathematical problem solving and achievement. This study investigated the genetic and environmental factors contributing to the observed differences in the anxiety people feel when confronted with mathematical tasks. In addition, the genetic and environmental mechanisms that link mathematical anxiety with math cognition and general anxiety were also explored. METHODS: Univariate and multivariate quantitative genetic models were conducted in a sample of 514 12-year-old twin siblings. RESULTS: Genetic factors accounted for roughly 40% of the variation in mathematical anxiety, with the remaining being accounted for by child-specific environmental factors. Multivariate genetic analyses suggested that mathematical anxiety was influenced by the genetic and nonfamilial environmental risk factors associated with general anxiety and additional independent genetic influences associated with math-based problem solving. CONCLUSIONS: The development of mathematical anxiety may involve not only exposure to negative experiences with mathematics, but also likely involves genetic risks related to both anxiety and math cognition. These results suggest that integrating cognitive and affective domains may be particularly important for mathematics and may extend to other areas of academic achievement.
Number sense in human infants
DOI:10.1111/j.1467-7687.2005.00395.x
URL
PMID:15647069
[本文引用: 1]
Four experiments used a preferential looking method to investigate 6-month-old infants' capacity to represent numerosity in visual-spatial displays. Building on previous findings that such infants discriminate between arrays of eight versus 16 discs, but not eight versus 12 discs (Xu & Spelke, 2000), Experiments 1 and 2 investigated whether infants' numerosity discrimination depends on the ratio of the two set sizes with even larger numerosities. Infants successfully discriminated between arrays of 16 versus 32 discs, but not 16 versus 24 discs, providing evidence that their discrimination shows the set-size ratio signature of numerosity discrimination in human adults, children and many non-human animals. Experiments 3 and 4 addressed a controversy concerning infants' ability to discriminate large numerosities (observed under conditions that control for total filled area, array size and density, item size and correlated properties such as brightness: Brannon, 2002; Xu, 2003b; Xu & Spelke, 2000) versus small numerosities (not observed under conditions that control for total contour length: Clearfield & Mix, 1999). To investigate the sources of these differing findings, Experiment 3 tested infants' large-number discrimination with controls for contour length, and Experiment 4 tested small-number discrimination with controls for total filled area. Infants successfully discriminated the large-number displays but showed no evidence of discriminating the small-number displays. These findings provide evidence that infants have robust abilities to represent large numerosities. In contrast, infants may fail to represent small numerosities in visual-spatial arrays with continuous quantity controls, consistent with the thesis that separate systems serve to represent large versus small numerosities.
The neurodevelopmental basis of math anxiety
DOI:10.1177/0956797611429134
URL
PMID:22434239
[本文引用: 1]
Math anxiety is a negative emotional reaction to situations involving mathematical problem solving. Math anxiety has a detrimental impact on an individual's long-term professional success, but its neurodevelopmental origins are unknown. In a functional MRI study on 7- to 9-year-old children, we showed that math anxiety was associated with hyperactivity in right amygdala regions that are important for processing negative emotions. In addition, we found that math anxiety was associated with reduced activity in posterior parietal and dorsolateral prefrontal cortex regions involved in mathematical reasoning. Multivariate classification analysis revealed distinct multivoxel activity patterns, which were independent of overall activation levels in the right amygdala. Furthermore, effective connectivity between the amygdala and ventromedial prefrontal cortex regions that regulate negative emotions was elevated in children with math anxiety. These effects were specific to math anxiety and unrelated to general anxiety, intelligence, working memory, or reading ability. Our study identified the neural correlates of math anxiety for the first time, and our findings have significant implications for its early identification and treatment.
Gamma-band oscillations in the primary somatosensory cortex—a direct and obligatory correlate of subjective pain intensity
DOI:10.1523/JNEUROSCI.5877-11.2012
URL
PMID:22649223
[本文引用: 1]
Electroencephalographic gamma band oscillations (GBOs) induced over the human primary somatosensory cortex (SI) by nociceptive stimuli have been hypothesized to reflect cortical processing involved directly in pain perception, because their magnitude correlates with pain intensity. However, as stimuli perceived as more painful are also more salient, an alternative interpretation of this correlation is that GBOs reflect unspecific stimulus-triggered attentional processing. In fact, this is suggested by recent observations that other features of the electroencephalographic (EEG) response correlate with pain perception when stimuli are presented in isolation, but not when their saliency is reduced by repetition. Here, by delivering trains of three nociceptive stimuli at a constant 1 s interval, and using different energies to elicit graded pain intensities, we demonstrate that GBOs recorded over SI always predict the subjective pain intensity, even when saliency is reduced by repetition. These results provide evidence for a close relationship between GBOs and the cortical activity subserving pain perception.
Visual perception can account for the close relation between numerosity processing and computational fluency
DOI:10.3389/fpsyg.2015.01364
URL
PMID:26441740
[本文引用: 1]
Studies have shown that numerosity processing (e.g., comparison of numbers of dots in two dot arrays) is significantly correlated with arithmetic performance. Researchers have attributed this association to the fact that both tasks share magnitude processing. The current investigation tested an alternative hypothesis, which states that visual perceptual ability (as measured by a figure-matching task) can account for the close relation between numerosity processing and arithmetic performance (computational fluency). Four hundred and twenty four third- to fifth-grade children (220 boys and 204 girls, 8.0-11.0 years old; 120 third graders, 146 fourth graders, and 158 fifth graders) were recruited from two schools (one urban and one suburban) in Beijing, China. Six classes were randomly selected from each school, and all students in each selected class participated in the study. All children were given a series of cognitive and mathematical tests, including numerosity comparison, figure matching, forward verbal working memory, visual tracing, non-verbal matrices reasoning, mental rotation, choice reaction time, arithmetic tests and curriculum-based mathematical achievement test. Results showed that figure-matching ability had higher correlations with numerosity processing and computational fluency than did other cognitive factors (e.g., forward verbal working memory, visual tracing, non-verbal matrix reasoning, mental rotation, and choice reaction time). More important, hierarchical multiple regression showed that figure matching ability accounted for the well-established association between numerosity processing and computational fluency. In support of the visual perception hypothesis, the results suggest that visual perceptual ability, rather than magnitude processing, may be the shared component of numerosity processing and arithmetic performance.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}