

1 引言
抑郁症是心境障碍的主要类型之一, 目前全球共有超过3亿名抑郁症患者, 其中每年约有80万人死于自杀(WHO, 2018)。患者对自我、世界及未来抱有负性态度, 对负性刺激过度关注(何振宏, 张丹丹, 罗跃嘉, 2015)。大量流行病学及神经生物学研究表明, 社会、生物及心理因素在抑郁症发生与发展的过程中共同起到了至关重要的作用(Kupferberg, Bicks, & Hasler, 2016)。研究发现, 社会孤立、更小的社交网络, 以及对社会支持的低感知度均与抑郁程度高度相关(Jacobson, Lord, & Newman, 2017)。加强对抑郁症社会功能障碍脑机制的研究, 并用以指导抑郁症的防治, 可预防心理和行为问题的发生, 具有重大现实意义。
合作是伴随人类发展的亲社会行为。与其他物种不同, 人类可以在没有血缘关系的情况下建立稳定的合作关系(Bowles & Gintis, 2011), 甚至在自己利益受损的情况下也是如此(Rilling & Sanfey, 2011)。这是因为合作行为会激活大脑的奖赏系统, 因此尽管有时物质奖赏减少了, 但人们会为了追求社会奖赏和自我肯定而进行合作(Ruff & Fehr, 2014)。囚徒困境游戏(prisoner’s dilemma game, PDG)是较常用的研究合作的范式。在囚徒困境游戏中, 两名玩家同时选择合作或背叛。对于每名玩家来说, 不论对家的决策如何, 选择背叛带来的个人收益高于选择合作。然而实验发现, 人们愿意频繁地尝试合作方案(Rilling et al., 2002), 大量PDG研究中观测到的合作率均值为50%左右(Camerer, 2003)。脑成像PDG研究发现, 合作激活了内侧前额叶皮质(medial prefrontal cortex, mPFC)、颞顶联合区(temporoparietal junction, TPJ)等心理理论相关脑区(Emonds, Declerck, Boone, Vandervliet, & Parizel, 2012), 眶额叶皮层(orbitofrontal cortex, OFC)、腹侧纹状体等奖赏加工脑区、以及背外侧前额叶(dorsolateral prefrontal cortex, dlPFC)等负责认知控制的脑区(Fermin et al., 2016)。而抑郁相关研究已表明, mPFC/OFC (Bludau et al., 2016)、TPJ (Poeppl et al., 2016)、dlPFC (Wise et al., 2017)在抑郁症患者或高抑郁倾向群体中的结构缩小、功能性激活不足、或与其他脑区的功能连接出现异常, 这可能是抑郁影响合作行为的神经机制之一。
目前已有少量研究考察了抑郁对合作的影响, 发现抑郁会降低个体的合作倾向(Kupferberg et al., 2016; Pulcu et al., 2015)。例如, Clark, Thorne, Hardy和Cropsey (2013)观察到, 抑郁症患者在PDG中难以维持互惠合作, 在公共物品游戏中比健康对照贡献的金额更少。Gradin等(2016)进行了此领域迄今唯一的一项脑成像研究, 他们采用功能磁共振技术在PDG中发现, 抑郁症患者在互惠合作中难以体验到社会奖赏带来的快感, 他们的伏隔核和背侧尾状核等奖赏脑区的激活显著低于健康对照; 同时患者的认知控制和情绪调节能力受损, 表现为其dlPFC激活减弱。此外, 由于合作为一项多人交互的社会行为, 抑郁个体在决策过程中是否决定合作还取决于同伴的特质。Sorgi和van 't Wout (2016)在解决这一问题上进行了很好的尝试, 他们在PDG中采用计算机模拟的假被试作为游戏对家, 发现高抑郁倾向被试虽然在与“无偏性对家” (即合作率为50%的假对家)进行游戏时比低抑郁倾向被试表现出更多的背叛行为, 但他们在与“高合作性对家” (即合作率为75%的假对家)进行游戏时却能表现出较稳定的合作行为。受此启发, 为了更直接地考察抑郁对合作的影响, 我们在本研究中将被试分为以下3组进行实验:低抑郁倾向-低抑郁倾向组, 高抑郁倾向-高抑郁倾向组, 高抑郁倾向-低抑郁倾向组。
本研究的神经活动观测手段选用了基于近红外光谱成像(near-infrared spectroscopy, NIRS)的超扫描技术(hyperscanning), 该技术同时记录并考察特定认知活动过程中两名或多名被试之间的大脑活动的同步性, 已成为社会认知神经科学的重要研究方法(Babiloni & Astolfi, 2014)。近年来, 研究者利用双人同步记录技术发现, 游戏双方在合作过程中mPFC、OFC、dlPFC以及右侧TPJ的含氧血红蛋白浓度序列或脑电周期性振荡的同步性增高(Abe et al., 2019; Cheng, Li, & Hu, 2015; Lu, Xue, Nozawa, & Hao, 2018; Xue, Lu, & Hao, 2018)。
综上, 本研究采用PDG范式, 按照抑郁得分将被试分为3组(低低抑郁组、高高抑郁组、高低抑郁组), 利用双人同步近红外脑成像技术考察抑郁倾向对合作的影响。根据上述与抑郁相关的研究结果, 本文假设:行为方面, 抑郁导致合作率降低(Clark et al., 2013; Pulcu et al., 2015), 情绪满意度降低(Gradin et al., 2016)。单人脑激活强度方面, 高抑郁倾向被试在mPFC (Bludau et al., 2016)、右侧TPJ (Poeppl et al., 2016)等心理理论相关脑区, OFC等奖赏加工脑区, 以及dlPFC (Gradin et al., 2016)等认知控制脑区的激活水平可能会低于低抑郁倾向被试。双人脑间同步性方面, 高抑郁倾向被试与对家的上述脑区的含氧血红蛋白浓度序列的同步性降低, 且当对家同样为高抑郁倾向被试时此现象更明显(Sorgi & van 't Wout, 2016)。
2 方法
2.1 被试
采用贝克抑郁量表(Beck Depression Inventory Second Edition, BDI-II)从3500名大学本科生中筛选被试。根据BDI-II常模, 本研究邀请分数 ≤ 13 (无抑郁)以及分数 ≥ 20 (中重度抑郁)的志愿者参与实验。由于实验中需将被试按照抑郁倾向高低随机分配至低-低组、高-高组、高-低组, 每种配对组在正式实验中拟搜集25对数据, 在预实验中拟搜集1对数据, 即每种配对组需26对被试参与实验, 3组共78对被试参与实验。因此, 本次实验纳入被试共156名, 其中高抑郁倾向(即BDI-II提示中重度抑郁)、低抑郁倾向(即BDI-II提示无抑郁)各78人。
正式实验在BDI-II筛查后的1周内进行。实验前所有被试填写抑郁自评量表(Self-Rating Depression Scale, SDS)和斯皮尔伯格特质焦虑量表(The Trait form of Spielberger's State-Trait Anxiety Inventory, STAI-T)。考虑到合作行为还可能受到焦虑的影响(Fett et al., 2016), 本研究匹配了高、低抑郁倾向组组间的特质焦虑水平, 以排除焦虑因素对结果的影响。根据SDS常模, 我们将被试分为两组:SDS < 0.5为低抑郁倾向组(n = 78), SDS ≥ 0.5为高抑郁倾向组(n = 78)。所有被试在事前BDI-II筛查和实验前SDS评分中获得的分组身份(即高、低抑郁倾向)均保持一致。两组被试在年龄、性别和STAI-T评分方面均无显著差异(表1)。被试均无癫痫、脑外伤等病史, 视力或矫正视力正常。所有被试均为右利手。共同完成实验的两名被试性别相同,互为陌生人, 在实验过程中不允许交流。实验方案经实验所在大学医学院伦理委员会批准。主试在实验前向每名被试解释了实验内容以及参加实验的收益和潜在风险。所有被试都自愿参加实验并签署了知情同意书。由于预实验进行顺利, 数据分析时将预实验的3对被试也纳入分析, 因此本研究在低-低组、高-高组、高-低组各有26对数据。
表1 高、低抑郁倾向被试的人口学特征(M ± SD)
| 变量 | 低抑郁倾向 (n = 78) | 高抑郁倾向 (n = 78) | 统计结果 |
|---|---|---|---|
| 年龄 | 20.4 ± 1.4 | 20.6 ± 1.6 | t(154) = -0.72, p = 0.470 |
| 性别, 男/女 | 40/38 | 39/39 | χ2(1) = 0.03, p = 0.873 |
| 抑郁自评量表(SDS) | 0.41 ± 0.06 | 0.55 ± 0.08 | t(154) = -12.7, p < 0.001 |
| 特质焦虑量表(STAI-T) | 21.7 ± 3.3 | 26.0 ± 3.5 | t(154) = -1.06, p = 0.291 |
2.2 实验过程
实验采用经典的PDG。在多轮游戏中, 两名玩家在合作和不合作之间进行选择, 每轮的收益取决于双人的选择。两名玩家的配对固定, 即在整个实验过程中不更换对家。如果双人均选择合作(cooperation-cooperation, CC), 则每人获得2元; 如果双人均选择不合作(defection-defection, DD), 每人获得1元; 如果单人选择合作(CD/DC), 则合作方得0元, 不合作方得3元。因此, 对个体而言, 选择不合作的预期收益高于选择合作, 但对双人而言, 合作会使两人的总收益大于不合作的情况(Gradin et al., 2016; Rilling et al., 2002)。
实验前, 主试向被试介绍游戏规则, 被试进行10个回合的练习。被试的最终收益正比于个人在任务中累积的总分。被试被鼓励最大限度地提高自己的收益。在实验中, 两名被试相对而坐, 每人各自面对一块计算机屏幕。实验共120个回合(试次), 分3个block进行, 每个block 8 min, block之间休息5 min。每一回合的时间流程如图1所示, 首先出现一个2×2的收益矩阵, 被试通过按键选择“合作”或“不合作”。1号被试按D和F键进行选择, 2号被试按J和K键进行选择。按键所对应的含义(合作/不合作)在不同的被试配对间进行平衡。当一名被试进行选择后, 他/她的编号会以红色高亮显示。在双人做出选择后, 收益矩阵中的对应单元从白色变为灰色。从收益矩阵开始出现到结果呈现结束共6 s。试次间的jitter为2~10 s的随机数。
图1
在PDG后, 被试需完成一份情绪问卷, 该问卷要求他们在4种结果下(双人合作CC、双人不合作DD、本人合作对家不合作CD、本人不合作对家合作DC)对以下5项内容进行9点评分:(1)收益的满意度, (2)对家的满意度, (3)高兴, (4)内疚, (5)愤怒(Gradin et al., 2016; Sun et al., 2016)。
2.3 近红外数据采集及通道定位
本研究使用NirScan-2442设备(丹阳慧创, 中国江苏)在任务中以连续波形式记录NIRS数据, 采样率21 Hz。每名被试的光极包含10个发射器(光源)和11个探测器(探头), 组成29个通道, 发射器和探测器的距离均值为3.2 cm (范围为2.8~3.6 cm)。光极的放置采用10/20系统脑电帽(EASYCAP, Herrsching, Germany)。基于已有研究(Cui, Bryant, & Reiss, 2012; Gradin et al., 2016), 本实验主要观测的脑区为前额叶和右侧颞顶联合区; 每名被试额叶有22个通道, 右侧颞顶联合区有7个通道(图2)。
图2

本文定义每个通道的中点(即发射器和探测器连线的中点)下方脑区为该通道的主要探测区域, 并以此为标准进行通道的脑区标定。通道中点的MNI坐标采用NFRI工具包计算(
2.4 近红外数据分析
数据分析使用Matlab R2017b (MathWorks, Natick, MA, USA)。按照以下步骤对信号进行预处理:(1)删除数据中光强过饱和的通道; (2)由光强信号计算得到光密度信号的变化值; (3)采用样条插值去除信号中的运动伪迹; (4)使用0.01~0.2 Hz的带通滤波器去除生理噪声及基线漂移; (5)基于修正的Beers-Lambert定律从光密度信号解算出HbO和Hb的浓度变化值(即Δ[HbO]和Δ[Hb])。由于Δ[HbO]比Δ[Hb]对条件间的变化更敏感, 后续分析仅使用Δ[HbO]数据。
本文所关注的脑区(regions of interest, ROI)分别对应以下brodmann分区:mPFC (8, 9, 10, 24, 32), OFC (10, 11, 47), dlPFC (8, 9, 10, 46), TPJ (39, 40)。参考通道定位(附表1)及光极排布(图2), 每个ROI对应的通道为:mPFC对应通道11、12、19、21, OFC对应通道1、2、4、5, 左侧dlPFC对应通道9、17、18, 右侧dlPFC对应通道16、20、22, 右侧TPJ 对应通道23、24、27、28。考虑到NIRS技术较低的空间分辨率以及较低的信噪比, 本文将脑激活水平以及脑间同步性指标先在每个ROI内进行通道间的平均后再进行统计分析(Dai et al., 2018)。
单人脑激活水平采用一般线性模型(general linear model, GLM)计算。以每个试次中结果反馈开始呈现的时刻为零时刻。在GLM中对β值进行估计时采用AR (n)模型(1 < n ≤ 30), 用极大似然法计算超定线性方程组的最小二乘解。在每个fNIRS通道中, 基于所有被试的被试内条件(CC、CD、DC和DD)的β值进行Fisher z变换, 求得归一化β值。统计时基于ROI求通道均值, 得到5个ROI的归一化β值。
双人脑间同步性以Pearson相关系数r作为度量指标(Dai et al., 2018)。首先计算每个通道3个被试内条件(CC、CD/DC和DD)双人Δ[HbO]的6 s时间序列之间的Pearson相关系数r, 再基于ROI求通道均值, 得到5个ROI的r值。经K-S正态检验, 确认r值符合正态分布, 无需进行Fisher z变换。
2.5 统计
统计分析采用SPSS Statistics 20.0 (IBM, Somers, USA)。除非有特别说明, 描述性统计量表示为“均值 ± 标准差”。对因变量进行多因素重复测量方差分析, 显著性水平为p < 0.05。单人脑激活强度的考察采用2(抑郁倾向:高、低) × 4(决策结果:CC、CD、DC和DD)的混合设计。双人脑间同步性的考察采用3(配对组:低-低抑郁倾向、高-高抑郁倾向、高-低抑郁倾向) × 3(决策结果:双人合作、双人不合作、单人合作)的混合设计。对每个ROI的ANOVA, 采用Greenhouse-Geisser方法对自由度进行校正, 采用Bonferroni方法进行事后多重比较矫正。Pearson相关系数r的显著性采用Bonferroni方法在条件间及ROI间进行多重比较矫正。
为了排除由“相似认知任务” (又称为条件相似性, condition similarity)而非“交互认知任务”引发的脑同步现象, 本研究使用置换检验(permutation test)考察脑间同步性的可靠程度。具体的做法是打乱真实的被试配对, 随机将没有真实互动的两名被试作为一个配对并计算脑间同步性指标, 这一过程重复500次, 可生成一个脑间同步性指标的零假设分布(即仅由条件相似性引起的脑间同步性改变)。如果我们发现的脑间同步性改变超出了该零假设分布的95%置信区间, 即可认为此脑间同步性改变更可能是由“交互认知任务”引发的脑同步现象。本文仅对这种脑同步效应进行报告和讨论。
3 结果
3.1 合作率和反应时
首先在单人层次上对合作行为进行分析。本文定义合作率为单个被试在任务中选“合作”的比例。所有被试的合作率为0.49 ± 0.12。在120个试次的任务中, 双方合作(CC) 33 ± 15试次, 单方合作(CD/DC) 51 ± 10试次, 双方背叛(DD) 35 ± 14试次。独立样本t检验结果表明, 高抑郁倾向被试的合作率(0.45 ± 0.12)显著低于低抑郁倾向被试的合作率(0.54 ± 0.10), t(154) = -4.89, p < 0.001。采用双因素(高/低抑郁倾向 × 合作/背叛)混合方差分析对反应时进行统计, 发现各条件间反应时无显著差异(F ≤ 2.64, p ≥ 0.106; 合作:高vs.低抑郁倾向 = 840 ± 326 ms vs. 912 ± 343 ms, 背叛:高vs.低抑郁倾向 = 839 ± 308 ms vs. 940 ± 408 ms)。
进一步地, 我们分析了对方前一次的决策结果对被试当前决策的影响。这种影响可能有4种模式:投桃报李(对方合作后我也合作, XCCX), 以德报怨(对方背叛后我还合作, XDCX), 以牙还牙(对方背叛后我也背叛, XDDX), 恩将仇报(对方合作后我却背叛, XCDX)。采用双因素(高/低抑郁倾向 × 4种决策模式)混合方差分析分别对4种决策模式的“出现概率”以及“反应时”两个因变量进行统计(描述性统计量见图3)。对于出现概率, 影响模式的主效应显著, F(3,462) = 59.1, p < 0.001, η2p = 0.277:投桃报李(0.31 ± 0.15)和以牙还牙(0.32 ± 0.12)的出现概率显著高于以德报怨(0.18 ± 0.07)和恩将仇报(0.19 ± 0.07; ps < 0.001)。高低抑郁组间效应以及二者的交互作用不显著(F ≤ 1.91, p ≥ 0.151)。对于反应时, 影响模式的主效应显著, F(3,462) = 4.72, p = 0.003, η2p = 0.030:投桃报李(843 ± 0.15 ms)的反应时显著短于以德报怨(899 ± 0.15 ms, p = 0.015)和恩将仇报(911 ± 0.15 ms, p = 0.005)。高低抑郁组间效应(F(1,154) = 2.37, p = 0.126)以及二者的交互作用不显著(F(3,462) = 2.02, p = 0.113)。
图3

图3
对方前一次的决策结果对被试当前决策的影响。A, 不同影响模式的出现概率; B, 不同影响模式对应的反应时。本研究关注的4种影响模式为:投桃报李(XCCX)、以德报怨(XDCX)、以牙还牙(XDDX)、恩将仇报(XCDX)。图中errorbar表示均值的标准误。
最后, 我们在双人层次上对合作行为进行分析。对互惠合作率(即CC的概率)进行单变量三水平统计, 发现组间效应显著, F(2,75) = 3.70, p = 0.029, η2p = 0.090。3组被试的互惠合作率有以下趋势:低-低抑郁倾向(0.32 ± 0.11) > 高-低抑郁倾向(0.28 ± 0.12) > 高-高抑郁倾向(0.23 ± 0.12); 其中低-低抑郁倾向组的互惠合作率显著大于高-高抑郁倾向组(p = 0.024), 高-低抑郁倾向组的互惠合作率与其他两组无显著差异(p ≥ 0.461)。
3.2 事后情绪评分
对5项情绪评分进行单人层面的双因素方差分析, 以下仅报告显著结果。
对收益的满意度方面, 决策结果的主效应显著, F(3,462) = 272, p < 0.001, η2p = 0.639:对家不合作时被试对收益的满意度降低, 即CD (3.00 ± 0.16)和DD (3.85 ± 0.14)条件小于CC (7.08 ± 0.12)和DC (7.32 ± 0.13)条件(ps < 0.001); 此外CD条件被试对收益的满意度低于DD条件(p < 0.001)。
对对家的满意度方面, 决策结果的主效应显著, F(3,462) = 184, p < 0.001, η2p = 0.545:对家不合作时被试对对家的满意度降低, 即CD (3.87 ± 0.16)和DD (4.39 ± 0.15)条件小于CC (7.57 ± 0.11)和DC (6.83 ± 0.14)条件(ps < 0.001); 此外CC条件被试对对家的满意度高于DC条件(p < 0.001)。抑郁倾向主效应显著, F(1,154) = 5.7, p = 0.018, η2p = 0.036:高抑郁倾向的被试对对家的满意度(5.55 ± 2.33)显著低于低抑郁倾向被试(5.98 ± 2.40)。
对自身高兴情绪评分方面, 决策结果的主效应显著, F(3,462) = 251, p < 0.001, η2p = 0.620:对家合作比对家不合作增加了被试的高兴程度。4个决策结果对应的高兴评分具有以下关系:CC (7.29 ± 1.44) > DC (6.54 ± 1.78) > DD (3.73 ± 1.85) > CD (2.96 ± 1.86), 两两间差异显著(ps ≤ 0.001)。
对自身内疚情绪评分方面, 决策结果的主效应显著, F(3,462) = 83.3, p < 0.001, η2p = 0.351:被试在DC情况下的内疚情绪最强(4.58 ± 2.45; ps < 0.001), 其次是DD (2.28 ± 1.87)情况, 被试在CC (1.68 ± 1.38; CC vs. DD, p = 0.004)和CD (1.89 ± 1.68; CC vs. DD, p = 0.063)情况下的内疚情绪最弱。
对自身愤怒情绪评分方面, 决策结果的主效应显著, F(3,462) = 96.0, p < 0.001, η2p = 0.384。4个决策结果对应的愤怒评分具有以下关系:CD (4.36 ± 2.54) > DD (2.72 ± 2.14) > DC (1.69 ± 1.36) > CC (1.36 ± 1.00), 两两间差异显著(ps ≤ 0.041)。
3.3 单人脑激活强度(归一化β值)
在OFC脑区, 决策结果主效应显著, F(3,462) = 10.6, p < 0.001, η2p = 0.065:CC条件的脑激活(0.219 ± 0.835)强于DC条件(0.159 ± 0.834, p < 0.001), 这二者的脑激活又强于CD (-0.216 ± 0.940, p ≤ 0.001)和DD条件(-0.167 ± 1.230, p ≤ 0.046), 后二者的差异不显著(p = 1.000)。组间效应和决策结果的交互作用显著, F(3,462) = 3.54, p = 0.043, η2p = 0.022; 图4A, 图5A。简单效应分析表明, 决策结果的效应(CC > DC > CD/DD)仅在低抑郁倾向组中显著(F(3,152) = 14.5, p < 0.001, η2p = 0.158; CC = 0.304 ± 0.811, CD = -0.359 ± 0.887, DC = 0.234 ± 0.811, DD = -0.306 ± 1.063), 在高抑郁倾向组中不显著(F < 1; CC = 0.133 ± 0.856, CD = -0.074 ± 0.974, DC = 0.083 ± 0.856, DD = -0.028 ± 1.368)。
图4

图4
单人脑激活强度的条件间差异。A, 抑郁倾向组间效应(高、低抑郁倾向)和决策结果(CC、CD、DC和DD)的交互作用; B, 抑郁倾向组间效应的主效应(高vs. 低抑郁倾向)。图中颜色代表方差分析的F值, 即颜色越红表示该脑区的交互作用或主效应对应的F值越大。
图5
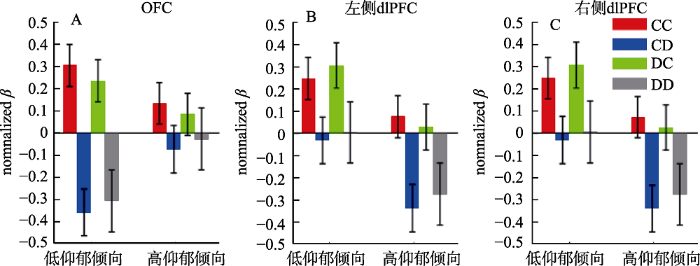
图5
不同条件下的脑激活水平(归一化β值)。A, OFC; B, 左侧dlPFC; C, 右侧dlPFC。4个条件为:双人合作CC、本人合作对家不合作CD、本人不合作对家合作DC、双人不合作DD)。图中errorbar表示均值的标准误。
未在mPFC和右侧TPJ脑区发现归一化β值在条件间的显著差异。
3.4 双人脑间同步性(r值)
在OFC脑区, 决策结果主效应显著, F(2,150) = 5.44, p = 0.005, η2p = 0.068 (图6A):双人合作条件的脑间相关性(0.193 ± 0.159)大于单人合作(0.135 ± 0.119, p = 0.021)和双人背叛条件(0.128 ± 0.173, p = 0.013), 后二者的差异不显著(p = 1.000)。组间效应和交互作用不显著(Fs ≤ 1.06, p ≥ 0.350)。
图6
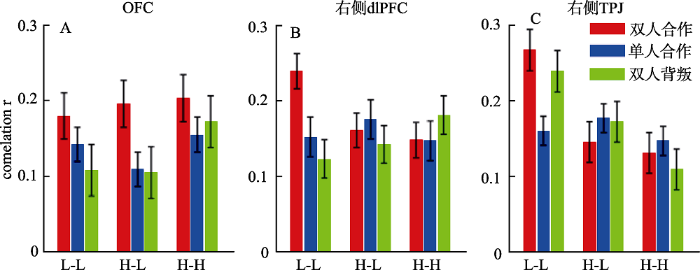
图6
不同条件下的脑间同步性。A, OFC; B, 右侧dlPFC; C, 右侧TPJ。抑郁倾向分组:L-L为低-低抑郁倾向组, H-L为高-低抑郁倾向组, H-H为高-高抑郁倾向组。本图中errorbar表示均值的标准误。
在右侧dlPFC脑区, 组间效应和决策结果的主效应不显著(Fs ≤ 1.99, p ≥ 0.142), 但二者的交互作用显著, F(4,150) = 3.62, p = 0.008, η2p = 0.088 (图6B)。简单效应分析表明, 低-低抑郁倾向组的脑间同步性在双人合作条件(0.239 ± 0.097)大于双人背叛(0.123 ± 0.107, p < 0.001)和单人合作条件(0.152 ± 0.151, p = 0.031), F(2,74) = 9.06, p < 0.001, η2p = 0.197, 但决策结果的效应在另两组中不显著(Fs < 1)。另一方向的简单效应分析表明, 双人合作条件下存在组间效应, F(2,75) = 4.40, p = 0.016, η2p = 0.105:低-低抑郁倾向组的脑间相关性大于高-高抑郁倾向组(p = 0.023)以及高-低抑郁倾向组(p = 0.066), 但单人合作和双人背叛条件下不存在组间效应(Fs ≤ 1.38, p ≥ 0.257)。
在右侧TPJ脑区, 相关系数r组间差异显著, F(2,75) = 7.74, p = 0.001, η2p = 0.171:低-低抑郁倾向组的脑间相关性(0.222 ± 0.116)大于高-高抑郁倾向组(0.129 ± 0.140, p = 0.001)以及高-低抑郁倾向组(0.165 ± 0.127, p = 0.057), 后二者的差异不显著(p = 0.414)。组间效应和决策结果的交互作用显著, F(4,150) = 3.50, p = 0.010, η2p = 0.085 (图6C)。简单效应分析表明, 组间效应仅在双人合作(F(2,75) = 7.49, p = 0.001, η2p = 0.167)和双人背叛条件下显著(F(2,75) = 5.69, p = 0.005, η2p = 0.132), 在单人合作条件下不显著(F < 1)。另一方向的简单效应分析表明, 低-低抑郁倾向组的脑间同步性在双人合作(0.267 ± 0.116, p = 0.007)和双人背叛条件(0.239 ± 0.101, p = 0.016)大于单人合作条件(0.160 ± 0.107), F(2,74) = 7.56, p = 0.001, η2p = 0.170, 但决策结果的效应在另两组中不显著(Fs < 1)。
未在左侧dlPFC和mFPC脑区发现相关系数r在条件间的显著差异。
3.5 脑活动指标对合作行为的预测
本部分考察脑活动指标是否可用于预测整个实验过程中被试的“合作率”和“互惠合作率”, 且该预测能力是否受到抑郁倾向的调节。
在单人层面, 多元回归分析以“合作率” (选择合作的概率)为因变量, 以5个ROI在被试选择合作条件下的归一化β值为预测变量, 结果发现回归模型不显著。单人层面的回归结果见表2。
表2 脑区激活(β值)对合作率的预测
| 分组模型 | 模型参数 | 标准化回归系数(B) | t值的显著性 (p) |
|---|---|---|---|
| 不分组 | R2 = 0.032 | OFC = -0.064 | 0.499 |
| (n = 156) | F(5,150) = 0.99 | TPJ = 0.096 | 0.275 |
| p = 0.428 | mPFC = -0.042 | 0.723 | |
| left dlPFC = -0.094 | 0.337 | ||
| right dlPFC = -0.042 | 0.641 | ||
| 低抑郁倾向 | R2 = 0.025 | OFC = -0.170 | 0.335 |
| (n = 78) | F(5,72) = 0.365 | TPJ = 0.085 | 0.486 |
| p = 0.871 | mPFC = 0.109 | 0.628 | |
| left dlPFC = -0.072 | 0.649 | ||
| right dlPFC = -0.003 | 0.982 | ||
| 高抑郁倾向 | R2 = 0.087 | OFC = -0.033 | 0.790 |
| (n = 78) | F(5,20) = 1.36 | TPJ = 0.191 | 0.169 |
| p = 0.248 | mPFC = -0.153 | 0.338 | |
| left dlPFC = -0.136 | 0.307 | ||
| right dlPFC = -0.122 | 0.337 |
在双人层面, 多元回归分析以“互惠合作率” (双人合作的概率)为因变量, 以5个ROI在互惠合作条件下的脑间同步性指标r为预测变量, 结果发现回归模型显著, 且OFC、右侧TPJ以及双侧dlPFC的脑间同步性均对互惠合作率的预测有贡献。进一步地, 分别建立在高-高、高-低、低-低抑郁倾向组中的回归模型, 发现仅低-低抑郁倾向组的回归模型显著:右侧TPJ和双侧dlPFC的脑间同步性能正向预测互惠合作率。该结果说明, 参与合作的两名被试的抑郁倾向是采用脑间同步性预测互惠合作率的调节变量。双人层面的回归结果见表3。
表3 脑间同步性(r值)对互惠合作率(CC%)的预测
| 分组模型 | 模型参数 | 标准化回归系数(B) | t值的显著性 (p) |
|---|---|---|---|
| 不分组 | R2 = 0.267 | OFC = 0.327 | 0.004 |
| (n = 78) | F(5,72) = 5.26 | TPJ = 0.270 | 0.020 |
| p < 0.001 | mPFC = 0.225 | 0.087 | |
| left dlPFC = 0.365 | 0.007 | ||
| right dlPFC = 0.387 | 0.003 | ||
| 低-低抑郁倾向 | R2 = 0.653 | OFC = 0.310 | 0.059 |
| (n = 26) | F(5,20) = 7.53 | TPJ = 0.440 | 0.008 |
| p < 0.001 | mPFC = 0.038 | 0.857 | |
| left dlPFC = 0.493 | 0.022 | ||
| right dlPFC = 0.874 | 0.000 | ||
| 高-低抑郁倾向 | R2 = 0.184 | OFC = 0.258 | 0.285 |
| (n = 26) | F(5,20) = 0.90 | TPJ = 0.277 | 0.280 |
| p = 0.499 | mPFC = 0.394 | 0.154 | |
| left dlPFC = 0.307 | 0.279 | ||
| right dlPFC = 0.060 | 0.825 | ||
| 高-高抑郁倾向 | R2 = 0.376 | OFC = 0.405 | 0.067 |
| (n = 26) | F(5,20) = 2.41 | TPJ = 0.205 | 0.345 |
| p = 0.073 | mPFC = 0.055 | 0.812 | |
| left dlPFC = 0.266 | 0.279 | ||
| right dlPFC = 0.162 | 0.493 |
4 讨论
4.1 基于行为数据的发现
本研究采用PDG考察了高低抑郁倾向人群在合作过程中的行为表现、情绪反应, 以及大脑皮层激活和脑间同步性。结果显示, 高抑郁倾向组比低抑郁倾向组的合作性更低, 与已有的抑郁症行为研究一致(Clark et al., 2013; Pulcu et al., 2015)。事后情绪评分结果表明, 高抑郁倾向被试对对家的满意度显著低于低抑郁倾向被试, 这与Gradin等(2016)在抑郁症患者PDG研究中的发现基本一致。有研究指出, 抑郁人群的这种在社会交往中表现出的“敌对”态度很可能是延长或加重他们抑郁症状的原因(Bird, Tarsia, & Schwannauer, 2018)。本文的贡献之一在于, 我们将真实互动的两名被试的抑郁倾向水平作为研究变量, 发现双人在PDG中的合作率有如下趋势:低-低抑郁倾向组 > 高-低抑郁倾向组 > 高-高抑郁倾向组。该发现与Sorgi和van 't Wout (2016)采用假被试进行的PDG研究结果吻合, 即高抑郁倾向被试在与高合作性对家(对应于本研究的低抑郁倾向被试)进行游戏时其合作意愿比与低合作性对家进行游戏时有所提高。我们的这一发现提示, 抑郁或抑郁倾向作为一种特质或个体差异变量, 它对合作的影响符合叠加原理, 即双人的合作率与参与合作的两个单人的合作意愿具有正相关关系。本文的研究结果可能也部分解释了Cheng等人(2015)的研究发现, 后者发现合作率在女性-女性组合中最低, 男性-男性组合中最高, 而男性-女性组合的合作率居中。这是因为众所周知, 女性更易患抑郁症, 即通常女性被试的抑郁倾向比男性高, 因此两名女性搭配进行实验任务时可能会比男男组合产生更低的合作率。
4.2 基于脑活动数据的发现
继Gradin等(2016)之后, 本研究是考察抑郁对合作影响的第二项脑成像研究。Gradin等(2016)在单人的PDG任务中(被试与电脑进行游戏)发现, 抑郁被试的dlPFC在合作任务中激活减弱, 本研究除了重复出该脑区的组间差异外, 还在右侧TPJ脑区发现两名高抑郁倾向被试的脑间同步性在任务中明显低于两名低抑郁倾向被试。这为抑郁倾向或抑郁症的评估又提供了一项潜在的神经学标记物。
具体的, 我们的脑激活数据显示, 高抑郁倾向被试双侧dlPFC的激活显著弱于低抑郁倾向被试。脑间同步性方面, 合作选项与抑郁倾向的交互作用显著, 仅在低-低抑郁倾向组能观察到互惠合作引起的右侧dlPFC脑间同步性增强, 这一结果与已有的研究一致, 即在合作情境下双侧(Lu et al., 2018)或右侧dlPFC (Cui et al., 2012; Xue et al., 2018)的脑间同步性增高。本研究的新发现在于, 我们观察到抑郁倾向对此同步性改变有明显的调节作用, 高抑郁倾向被试参与的合作游戏不再能观察到互惠合作引起的脑间同步性增强。
dlPFC是主要的认知控制脑区(Miller & Cohen, 2001)。PDG研究发现, 成功的合作需要dlPFC的参与(Fermin et al., 2016), 采用TMS抑制dlPFC活动水平会导致合作行为减少(Soutschek, Sauter, & Schubert, 2015)。目前认为dlPFC在PDG中的作用是抑制“自私冲动”, 控制和调节自我利益与双人共同利益之间的冲突(Grecucci, Giorgetta, van’t Wout, Bonini, & Sanfey, 2013)。dlPFC脑区欠激活会导致认知控制和情绪调节异常(Gradin et al., 2016; Rilling & Sanfey, 2011)。例如, Knoch, Pascual-Leone, Meyer, Treyer和Fehr (2006)在最后通牒游戏中采用TMS抑制“方案接受者”的右侧dlPFC, 发现他们对分配方案的拒绝率下降, 这说明接受者在dlPFC功能损毁后更看重己方的短暂利益。类似的, Strang等(2015)在独裁者游戏中采用TMS抑制“方案分配者”的右侧dlPFC, 导致后者做出了更多的自私分配。
dlPFC是抑郁症患者脑异常最为突出的一个脑区, 也是目前临床采用TMS治疗难治性抑郁症的靶点脑区。dlPFC在抑郁人群中的结构和功能异常通常被发现位于双侧、左侧或右侧。例如, 有研究者认为杏仁核与双侧dlPFC等脑区的连接减弱是抑郁症状的核心病理生理学因素(Satterthwaite et al., 2016); 与健康对照相比, 抑郁症患者双侧dlPFC的灰质体积下降, 且dlPFC体积与抑郁反刍(rumination)的程度成反比(Wang et al., 2015)。左侧dlPFC异常的证据例如:抑郁症患者在PDG中左侧dlPFC的激活显著低于健康对照(Gradin et al., 2016); 左侧dlPFC与其他脑区的功能连接可用于预测患者接收TMS治疗的疗效(Richieri et al., 2018)。右侧dlPFC异常的证据例如:右侧dlPFC灰质体积缩小是抑郁症不同于双相障碍等其他情绪障碍的特异性大脑结构改变(Wise et al., 2017); 患者“认知控制脑网络”内部的功能连接减弱, 尤其是右侧dlPFC和右侧顶下小叶的 功能连接减弱(Stange et al., 2017)。本研究发现, 高抑郁倾向被试在整个PDG过程中双侧dlPFC的激活弱于低抑郁倾向被试。基于本文以及上述抑郁人群dlPFC的研究结果, 我们认为dlPFC的异常子区域(双侧、左侧、右侧)可能会由于抑郁严重程度、实验范式及考察变量的不同而改变。考虑到目前临床针对dlPFC的TMS治疗方案几乎均瞄准患者的左侧dlPFC (Kar, 2019; Sonmez et al., 2019), 我们建议未来的治疗可以结合个体的抑郁症状以及需要重点改善的认知功能, 灵活地选择双侧、左侧或右侧dlPFC予以治疗。例如, 本研究结果提示, 同时激活患者双侧dlPFC可能比仅激活左侧dlPFC更有利于增加患者在社会决策过程中的合作行为。
本研究在Gradin等(2016)的单人PGD研究基础上新发现了右侧TPJ脑区在条件间的改变, 即观察到该脑区的脑间同步性受到抑郁倾向主效应、合作选项与抑郁倾向交互作用的影响:低-低抑郁倾向组的右侧TPJ脑间同步性高于高-高以及高-低抑郁倾向组, 且这一效应当且仅当参与PDG双方的选择相同时(CC或DD条件)显著。TPJ (特别是右侧TPJ)是心理理论的重要脑区, 在注意力转移和推测他人意图过程中发挥着重要作用(Molenberghs, Johnson, Henry, & Mattingley, 2016)。抑郁症患者心理理论能力下降(Bora & Berk, 2016), 右侧TPJ与其他脑区的功能连接异常(Poeppl et al., 2016), TPJ静息态的激活水平正比于患者接受治疗后的疗效(Kraus et al., 2019)。本文发现的TPJ和dlPFC的脑间同步性结果具有一定的相似性, 即抑郁对合作选择在脑间同步性上的效应具有调节作用。这两个脑区的区别是, 在低-低抑郁倾向组可以观察到dlPFC仅在CC条件下脑间同步性增强, 而TPJ在CC和DD条件下脑间同步性均有增强。此前在双人合作领域开展的脑同步性研究几乎均采用了block设计, 这些研究结果表明, 与竞争情境或单人任务相比, 双人在合作block的TPJ脑间同步性增高(Abe et al., 2019; Lu et al., 2018; Xue et al., 2018)。此外, Gradin等(2016)虽采用PDG范式考察了抑郁症患者的脑激活异常, 却没有考察脑间同步性指标。本研究首次利用脑成像技术考察两名被试在PDG中的脑间同步性, 发现低-低抑郁倾向组不仅在互惠合作条件而且在双人背叛条件, 都具有显著上升的TPJ脑间同步性。这提示在PDG的双人背叛条件TPJ脑区也参与了“揣测他人心理”的认知过程:两名被试在结果呈现前均猜测或希望对方会选择合作选项, 在结果呈现后发现预期错误, 又调用心理理论脑区即TPJ重新评估对家的合作意愿。
本研究关注的另一个脑区是OFC。结果显示, OFC在互惠合作条件(CC)激活最显著, 己方背叛对方合作时(DC)激活次之, 对方背叛时(CD/DD)激活最弱。OFC是公认的奖赏加工重要脑区(Kringelbach, O'Doherty, Rolls, & Andrews, 2003)。合作是常见的社会奖赏之一, 合作行为可以显著激活OFC、伏隔核、尾状核等奖赏加工脑区(Rilling et al., 2002)。与我们的结果类似, Sun等(2016)利用PDG范式研究发现, 互惠合作比其他3种情况(CD/DC/ DD)更显著地激活了OFC和腹侧纹状体。本研究进一步发现OFC的激活在CC条件(个人收益2元)强于DC条件(个人收益3元), 这提示相比于金钱奖赏, OFC可能对社会奖赏更敏感。在双人脑同步性方面, 本研究发现互惠合作条件下OFC脑间同步性显著强于单人合作和双人背叛条件, 该结果与Cheng等(2015)在合作按键任务中的发现一致, 即OFC脑间同步性在合作情境下显著提高。
更重要的是, 本研究在OFC脑区发现了决策结果与抑郁倾向的交互作用, 即OFC在奖赏条件下(CC/DC)的显著激活仅在低抑郁倾向组中存在。已有研究表明, OFC脑区异常是抑郁症的重要神经机制。研究发现, 患者或抑郁倾向人群的OFC灰质体积缩小(Saleh et al., 2017), OFC与其他脑区的功能连接强度减弱(Frodl et al., 2010)。与本文的发现类似, Rilling等(2007)在人格障碍(psychopathy)群体的PDG研究中也发现, 与低人格障碍组相比, 高人格障碍组更多地选择背叛、难以维持互惠合作, 且在合作过程中其OFC的激活更弱。值得一提的是, 本文发现的决策结果与抑郁倾向的交互作用主要体现在内侧OFC区域(medial OFC; 参见图3A), 这一结果与目前学者们对OFC功能分区的认识是一致的, 即内侧OFC与奖赏加工相关, 主要负责奖赏价值的计算、奖赏期待以及奖赏预期误差的加工, 而外侧OFC与非奖赏加工或损失加工有关(Grabenhorst & Rolls, 2011)。与本文的发现相似, Rothkirch, Tonn, Köhler和Sterzer (2017)在一项金钱奖赏/损失任务中观察到, 抑郁症患者内侧OFC的激活明显弱于健康对照组, 且内侧OFC和腹侧纹状体的激活水平反比于患者的快感缺失程度; Cheng等(2018)发现患者内侧OFC与杏仁核的静息态功能连接减弱, 且减弱的程度与抑郁量表得分以及抑郁病程的长度显著相关; Cheng等(2016)通过对大样本的抑郁症患者静息态数据分析发现, 患者的内侧OFC与海马旁回、颞中回等记忆相关脑区的功能连接减弱, 且内侧OFC与颞中回连接强度减弱的程度正比于抑郁量表得分。这些研究结果(包括本文的发现)一致性地说明, 抑郁症患者或高抑郁倾向人群的内侧OFC在奖赏加工特别是社会性奖赏加工过程中激活不足或功能性连接下降。
4.3 脑间同步性对互惠合作的预测
通过回归分析我们发现, 在整个样本中, OFC、右侧TPJ以及双侧dlPFC的脑间同步性指标能预测互惠合作率。这是在利用相同范式进行单人实验(即被试与电脑互动)或假被试实验时(即被试与假被试互动)无法获得的信息, 提示双人或多人同步记录可以在传统的单人指标之外, 提供有潜在应用价值的、反映人际行为的脑活动参数。我们还尝试了采用个体脑区的激活水平去预测个体的合作率, 但回归模型并不显著, 这可能是因为合作涉及两个或两个以上个体, 仅用单人的脑激活强度去评估合作行为时所能利用的信息量太小。与本文类似, 另一些研究也发现脑间同步性指标可以对社会认知或交流行为进行预测。例如Jiang等(2015)考察了自由谈话过程中“领导者”角色的出现过程, 发现利用领导者-非领导者以及非领导者-非领导者之间的TPJ脑间同步性指标差异, 可以准确预测出谈话中出现的领导者。又如, Liu等(2019)的研究发现, 在有先验知识的面对面的教学过程中, 师生左侧前额叶的脑间同步性指标可以预测教学效果。以上这些结果均表明了脑间同步性指标对研究社会认知活动的重要性, 同时也提示了hyperscanning技术在神经经济学、管理学、教育学、市场营销等领域中推广应用的潜在价值。更重要的是, 本研究还发现双人的抑郁倾向是脑间同步性对合作行为预测的调节变量, 这提示抑郁倾向会对互惠合作产生显著的影响。
需要指出的是, 由于数据量的限制, 本研究实际仅考察了脑间同步性指标与互惠合作率的相关关系。下一步需借助机器学习算法并基于相邻试次的数据(例如Zhang, Lin, Jing, Feng, & Gu, 2019)或相邻block的数据(例如Reindl, Gerloff, Scharke, & Konrad, 2018)对社会行为进行实质性的预测。
4.4 研究局限性
本研究存在两点局限。第一, 我们所纳入的被试大部分未达到抑郁症的临床诊断标准, 因此将本研究结果推广至抑郁症人群时应持谨慎态度。第二, 本研究采用的fNIRS技术空间分辨率不足, 且各探测通道的定位可能存在一定个体差异。因此在数据分析时我们使用了ROI, 不在特定的皮层沟回考察成像结果。第三, fNIRS技术仅能检测到大脑皮层2~3 cm深度的血氧浓度改变, 无法探测一些与合作相关的深部脑区, 例如纹状体、脑岛等。这可能也是本文在mPFC脑区未发现阳性结果的原因。
5 结论
本研究采用PDG范式考察了高低抑郁倾向人群在合作过程中的行为表现、情绪反应, 以及大脑皮层激活水平和脑间同步性。行为和情绪自评结果显示, 高抑郁倾向被试比低抑郁倾被试的合作率更低, 对对家的满意度也更低。脑成像结果显示, 第一, OFC在互惠合作条件下激活最显著, 在己方背叛对方合作时激活次之, 且在此两种条件下的显著性激活仅在低抑郁倾向被试中才能观察到。这说明高抑郁倾向人群在奖赏特别是社会性奖赏加工中存在缺陷。第二, 高抑郁倾向被试双侧dlPFC的激活显著弱于低抑郁倾向被试, 抑郁对右侧dlPFC脑间同步性有调节作用, 在高抑郁倾向被试中不再能观察到互惠合作引起的脑间同步性增强。第三, 低-低抑郁倾向组的右侧TPJ脑间同步性高于高-高以及高-低抑郁倾向组, 且这一效应当且仅当参与PDG的双方选择相同时(均合作或均背叛)显著。上述结果说明, 高抑郁倾向群体在社会奖赏加工(OFC)、个人与集体利益的冲突控制(dlPFC)以及心理理论(TPJ)三项与合作相关的认知功能中均存在缺陷。
附录:
附表1 近红外通道的空间定位
| 通道编号 | MNI坐标 | 通道起止 | Brodmann模板 (脑区占通道的百分比)* | LPBA40模板 (脑区占通道的百分比)* |
|---|---|---|---|---|
| 1 | -34, 63, -8 | Fp1-AF7 | 10 - Frontopolar area (0.70) | L middle frontal gyrus (0.74) |
| 2 | -12, 71, -5 | Fp1-Fpz | 10 - Frontopolar area (0.80) | L superior frontal gyrus (0.93) |
| 3 | -23, 68, 2 | Fp1-AF3 | 10 - Frontopolar area (1) | L middle frontal gyrus (0.97) |
| 4 | 14, 71, -5 | Fp2-Fpz | 10 - Frontopolar area (0.88) | R middle frontal gyrus (1) |
| 5 | 36, 64, -9 | Fp2-AF8 | 10 - Frontopolar area (0.72) | R inferior frontal gyrus (0.62) |
| 6 | 26, 68, 2 | Fp2-AF4 | 10 - Frontopolar area (1) | R middle frontal gyrus (1) |
| 7 | -46, 51, 1 | F5-AF7 | 10 - Frontopolar area (0.53) | L inferior frontal gyrus (0.92) |
| 8 | -41, 55, 16 | F5-AF3 | 10 - Frontopolar area (0.85) | L middle frontal gyrus (1) |
| 通道编号 | MNI坐标 | 通道起止 | Brodmann模板 (脑区占通道的百分比)* | LPBA40模板 (脑区占通道的百分比)* |
| 9 | -48, 35, 25 | F5-FFC3 | 9/46 - Dorsolateral prefrontal cortex (0.86) | L middle frontal gyrus (0.60) |
| 10 | 2, 69, 11 | AFz-Fpz | 10 - Frontopolar area (1) | R superior frontal gyrus (0.74) |
| 11 | -15, 66, 23 | AFz-AF3 | 10 - Frontopolar area (1) | L middle frontal gyrus (0.50) |
| 12 | 17, 67, 24 | AFz-AF4 | 10 - Frontopolar area (1) | R middle frontal gyrus (0.96) |
| 13 | 2, 56, 38 | AFz-Fz | 9 - Dorsolateral prefrontal cortex (0.96) | R superior frontal gyrus (0.85) |
| 14 | 48, 51, 2 | F6-AF8 | 47 - Inferior prefrontal gyrus (0.47) | R inferior frontal gyrus (0.96) |
| 15 | 43, 55, 16 | F6-AF4 | 10 - Frontopolar area (0.93) | R middle frontal gyrus (0.65) |
| 16 | 50, 35, 26 | F6-FFC4 | 9/46 - Dorsolateral prefrontal cortex (0.86) | R middle frontal gyrus (0.60) |
| 17 | -26, 56, 30 | F1-AF3 | 10 - Frontopolar area (0.54) | L middle frontal gyrus (1) |
| 18 | -33, 38, 43 | F1-FFC3 | 8 - Includes Frontal eye fields (0.51) | L middle frontal gyrus (0.60) |
| 19 | -10, 45, 51 | F1-Fz | 8 - Includes Frontal eye fields (1) | L superior frontal gyrus (1) |
| 20 | 29, 56, 31 | F2-AF3 | 9 - Dorsolateral prefrontal cortex (0.53) | R middle frontal gyrus (1) |
| 21 | 13, 45, 51 | F2-Fz | 8 - Includes Frontal eye fields (1) | R superior frontal gyrus (0.98) |
| 22 | 35, 38, 44 | F2-FFC4 | 8 - Includes Frontal eye fields (0.57) | R middle frontal gyrus (0.72) |
| 23 | 62, -43, 45 | CP4-CP6 | 40 - Supramarginal gyrus (1) | R supramarginal gyrus (0.52) |
| 24 | 50, -56, 53 | CP4-P4 | 40 - Supramarginal gyrus (1) | R angular gyrus (1) |
| 25 | 69, -43, 10 | TP8-CP6 | 22 - Superior Temporal Gyrus (1) | R middle temporal gyrus (0.85) |
| 26 | 64, -56, 12 | TP8-P8 | 37 - Fusiform gyrus (0.91) | R middle temporal gyrus (0.93) |
| 27 | 62, -56, 29 | P6-CP6 | 40 - Supramarginal gyrus (0.71) | R angular gyrus (1) |
| 28 | 51, -68, 40 | P6-P4 | 39 - Angular gyrus (1) | R angular gyrus (1) |
| 29 | 57, -67, 13 | P6-P8 | 39 - Angular gyrus (0.56) | R middle occipital gyrus (0.92) |
注:*每条近红外通道可能覆盖多个脑区。为节省空间, 此处仅列出与近红外通道重合度最大的脑区。
参考文献
Neural correlates of online cooperation during joint force production
DOI:10.1016/j.neuroimage.2019.02.003
URL
PMID:30739061
[本文引用: 2]

During joint action, two or more persons depend on each other to accomplish a goal. This mutual recursion, or circular dependency, is one of the characteristics of cooperation. To evaluate the neural substrates of cooperation, we conducted a hyperscanning functional MRI study in which 19 dyads performed a joint force-production task. The goal of the task was to match their average grip forces to the target value (20% of their maximum grip forces) through visual feedback over a 30-s period; the task required taking into account other-produced force to regulate the self-generated one in real time, which represented cooperation. Time-series data of the dyad's exerted grip forces were recorded, and the noise contribution ratio (NCR), a measure of influence from the partner, was computed using a multivariate autoregressive model to identify the degree to which each participant's grip force was explained by that of their partner's, i.e., the degree of cooperation. Compared with the single force-production task, the joint task enhanced the NCR and activated the mentalizing system, including the medial prefrontal cortex, precuneus, and bilateral posterior subdivision of the temporoparietal junction (TPJ). In addition, specific activation of the anterior subdivision of the right TPJ significantly and positively correlated with the NCR across participants during the joint task. The effective connectivity of the anterior to posterior TPJ was upregulated when participants coordinated their grip forces. Finally, the joint task enhanced cross-brain functional connectivity of the right anterior TPJ, indicating shared attention toward the temporal patterns of the motor output of the partner. Since the posterior TPJ is part of the mentalizing system for tracking the intention of perceived agents, our findings indicate that cooperation, i.e., the degree of adjustment of individual motor output depending on that of the partner, is mediated by the interconnected subdivisions of the right TPJ.
Social neuroscience and hyperscanning techniques: Past, present and future
DOI:10.1016/j.neubiorev.2012.07.006
URL
PMID:22917915
[本文引用: 1]
This paper reviews the published literature on the hyperscanning methodologies using hemodynamic or neuro-electric modalities. In particular, we describe how different brain recording devices have been employed in different experimental paradigms to gain information about the subtle nature of human interactions. This review also included papers based on single-subject recordings in which a correlation was found between the activities of different (non-simultaneously recorded) participants in the experiment. The descriptions begin with the methodological issues related to the simultaneous measurements and the descriptions of the results generated by such approaches will follow. Finally, a discussion of the possible future uses of such new approaches to explore human social interactions will be presented.
Interpersonal styles in major and chronic depression: A systematic review and meta-analysis
DOI:10.1016/j.jad.2018.05.057
URL
PMID:29990668
[本文引用: 1]
McCullough's (2000) theory of chronic depression posits that a hostile-submissive interpersonal style distinguishes chronically depressed individuals from those with Major Depressive Disorder (MDD). This study sought to determine to what extent hostility and submissiveness feature in MDD, and whether there is evidence for a stronger effect in chronic depression.
Medial prefrontal aberrations in major depressive disorder revealed by cytoarchitectonically informed voxel-based morphometry
DOI:10.1176/appi.ajp.2015.15030349
URL
PMID:26621569
[本文引用: 2]
The heterogeneous human frontal pole has been identified as a node in the dysfunctional network of major depressive disorder. The contribution of the medial (socio-affective) versus lateral (cognitive) frontal pole to major depression pathogenesis is currently unclear. The authors performed morphometric comparison of the microstructurally informed subdivisions of human frontal pole between depressed patients and comparison subjects using both uni- and multivariate statistics.
Theory of mind in major depressive disorder: A meta-analysis
DOI:10.1016/j.jad.2015.11.023
URL
PMID:26655114
[本文引用: 1]
Social cognitive deficits can contribute to risk for depression and to psychosocial impairment during depression. However, available evidence suggests that emotion recognition is only marginally impaired in major depressive disorder (MDD). Recent studies have investigated theory of mind (ToM) abilities, a cognitively more demanding aspect of social cognition.
Medial reward and lateral non-reward orbitofrontal cortex circuits change in opposite directions in depression
DOI:10.1093/brain/aww255
URL
PMID:27742666
[本文引用: 1]
The first brain-wide voxel-level resting state functional connectivity neuroimaging analysis of depression is reported, with 421 patients with major depressive disorder and 488 control subjects. Resting state functional connectivity between different voxels reflects correlations of activity between those voxels and is a fundamental tool in helping to understand the brain regions with altered connectivity and function in depression. One major circuit with altered functional connectivity involved the medial orbitofrontal cortex Brodmann area 13, which is implicated in reward, and which had reduced functional connectivity in depression with memory systems in the parahippocampal gyrus and medial temporal lobe, especially involving the perirhinal cortex Brodmann area 36 and entorhinal cortex Brodmann area 28. The Hamilton Depression Rating Scale scores were correlated with weakened functional connectivity of the medial orbitofrontal cortex Brodmann area 13. Thus in depression there is decreased reward-related and memory system functional connectivity, and this is related to the depressed symptoms. The lateral orbitofrontal cortex Brodmann area 47/12, involved in non-reward and punishing events, did not have this reduced functional connectivity with memory systems. Second, the lateral orbitofrontal cortex Brodmann area 47/12 had increased functional connectivity with the precuneus, the angular gyrus, and the temporal visual cortex Brodmann area 21. This enhanced functional connectivity of the non-reward/punishment system (Brodmann area 47/12) with the precuneus (involved in the sense of self and agency), and the angular gyrus (involved in language) is thus related to the explicit affectively negative sense of the self, and of self-esteem, in depression. A comparison of the functional connectivity in 185 depressed patients not receiving medication and 182 patients receiving medication showed that the functional connectivity of the lateral orbitofrontal cortex Brodmann area 47/12 with these three brain areas was lower in the medicated than the unmedicated patients. This is consistent with the hypothesis that the increased functional connectivity of the lateral orbitofrontal cortex Brodmann area 47/12 is related to depression. Relating the changes in cortical connectivity to our understanding of the functions of different parts of the orbitofrontal cortex in emotion helps to provide new insight into the brain changes related to depression.
Functional connectivity of the human amygdala in health and in depression
DOI:10.1093/scan/nsy032
URL
PMID:29767786
[本文引用: 1]
To analyse the functioning of the amygdala in depression, we performed the first voxel-level resting state functional-connectivity neuroimaging analysis of depression of voxels in the amygdala with all other voxels in the brain, with 336 patients with major depressive disorder and 350 controls. Amygdala voxels had decreased functional connectivity (FC) with the orbitofrontal cortex, temporal lobe areas, including the temporal pole, inferior temporal gyrus and the parahippocampal gyrus. The reductions in the strengths of the FC of the amygdala voxels with the medial orbitofrontal cortex and temporal lobe voxels were correlated with increases in the Beck Depression Inventory score and in the duration of illness measures of depression. Parcellation analysis in 350 healthy controls based on voxel-level FC showed that the basal division of the amygdala has high FC with medial orbitofrontal cortex areas, and the dorsolateral amygdala has strong FC with the lateral orbitofrontal cortex and related ventral parts of the inferior frontal gyrus. In depression, the basal amygdala division had especially reduced FC with the medial orbitofrontal cortex, which is involved in reward; and the dorsolateral amygdala subdivision had relatively reduced FC with the lateral orbitofrontal cortex, which is involved in non-reward.
Synchronous brain activity during cooperative exchange depends on gender of partner: A fNIRS-based hyperscanning study
DOI:10.1002/hbm.22754
URL
PMID:25691124
[本文引用: 5]
Previous studies have shown that brain activity between partners is synchronized during cooperative exchange. Whether this neural synchronization depends on the gender of partner (i.e., opposite or same to the participant) is open to be explored. In current study, we used functional near-infrared spectroscopy (fNIRS) based hyperscanning to study cooperation in a two-person game (female-female, female-male, and male-male) while assaying brain-to-brain interactions. Cooperation was greater in male-male pairs than in female-female pairs, with intermediate cooperation levels for female-male pairs. More importantly, in dyads with partners with opposite gender (female-male pairs), we found significant task-related cross-brain coherence in frontal regions (i.e., frontopolar cortex, orbitofrontal cortex, and left dorsolateral prefrontal cortex) whereas the cooperation in same gender dyads (female-female pairs and male-male pairs) was not associated with such synchronization. Moreover, the changes of such interbrain coherence across task blocks were significantly correlated with change in degree of cooperation only in mixed-sex dyads. These findings suggested that different neural processes underlie cooperation between mixed-sex and same-sex dyadic interactions.
Cooperation and depressive symptoms
DOI:10.1016/j.jad.2013.05.011
URL
PMID:23726777
[本文引用: 2]
Deficits in pro-social cooperation are common in many individuals with mental illnesses such as depression. For decades, researchers have used economic game paradigms to compare cross-cultural cooperative behavior. However, research using economic games to assess cooperative behavior in clinical populations is in the early stages. We hypothesized that individuals with greater depressive symptoms would struggle to maintain reciprocity in iterative games, but not in single-iteration games measuring personal values.
NIRS-based hyperscanning reveals increased interpersonal coherence in superior frontal cortex during cooperation
DOI:10.1016/j.neuroimage.2011.09.003
URL
PMID:21933717
[本文引用: 3]
We used Near-Infrared Spectroscopy (NIRS) to simultaneously measure brain activity in two people while they played a computer-based cooperation game side by side. Inter-brain activity coherence was calculated between the two participants. We found that the coherence between signals generated by participants' right superior frontal cortices increased during cooperation, but not during competition. Increased coherence was also associated with better cooperation performance. To our knowledge, this work represents the first use of a single NIRS instrument for simultaneous measurements of brain activity in two people. This study demonstrates the use of NIRS-based hyperscanning in studies of social interaction in a naturalistic environment.
Holistic cognitive and neural processes: a fNIRS-hyperscanning study on interpersonal sensorimotor synchronization
DOI:10.1093/scan/nsy090
URL
PMID:30321411
Interpersonal sensorimotor synchronization (interpersonal SMS) is the foundation of complex human social interaction. Previous studies primarily focused on the individual cognitive processes of interpersonal SMS. However, all individuals compose an entire interaction system with emerged holistic properties during interpersonal SMS. Therefore, we proposed the `holistic cognitive and neural processes' of interpersonal SMS and defined quantitative measurements that included Holistic Correction Gain (HCG), Holistic Timekeeper Variance (HTV) and Holistic Motor Variance (HMV) based on linear error correction model and inter-brain couplings obtained by hyperscanning technique. We performed a joint-tapping experiment including bidirectional and unidirectional conditions using functional near-infrared spectroscopy (fNIRS) hyperscanning to evaluate effects of these holistic processes on synchronization performance. We found that the dyads' performance highly correlated with the integrated effect of holistic cognitive processes in both conditions. Each holistic cognitive process played different roles in interpersonal SMS. HCG was critical to maintain synchronization. HTV related to mentalizing others' behavior. Holistic neural process, the inter-brain coupling of right prefrontal cortex (PFC), was significantly different between bidirectional and unidirectional conditions, which suggested the existence of neural markers at holistic level in interpersonal SMS.
The cognitive demands on cooperation in social dilemmas: An fMRI study
DOI:10.1080/17470919.2012.655426
URL
PMID:22293039
[本文引用: 3]
This study uses fMRI to investigate the cognitive demands of decision-making in two types of cooperation games: a prisoner's dilemma (PD) eliciting a temptation to free-ride, leading to a dominant, self-interested response, and a stag hunt (SH) that has no dominant response but offers pay-off incentives that make mutual cooperation collectively beneficial but risky. Consequently, the PD poses greater conflict between self- and collective interest, greater demands for computational reasoning to derive the optimal solution, and greater demands for mentalizing to infer the intentions of others. Consistent with these differences between the two games, the results indicate that the PD is associated with increased activity in the anterior cingulate gyrus, prefrontal cortex, parietal lobe, and temporoparietal junction. With less conflict, the demands for computation and mentalizing are reduced in the SH, and cooperation levels increase dramatically. The differences in brain activation elicited by the different incentive structures of the PD and the SH appear to be independent of individual differences in revealed social preferences.
Representation of economic preferences in the structure and function of the amygdala and prefrontal cortex
DOI:10.1038/srep20982
URL
PMID:26876988
[本文引用: 1]
Social value orientations (SVOs) are economic preferences for the distribution of resources - prosocial individuals are more cooperative and egalitarian than are proselfs. Despite the social and economic implications of SVOs, no systematic studies have examined their neural correlates. We investigated the amygdala and dorsolateral prefrontal cortex (DLPFC) structures and functions in prosocials and proselfs by functional magnetic resonance imaging and evaluated cooperative behavior in the Prisoner's Dilemma game. We found for the first time that amygdala volume was larger in prosocials and positively correlated with cooperation, while DLPFC volume was larger in proselfs and negatively correlated with cooperation. Proselfs' decisions were marked by strong DLPFC and weak amygdala activity, and prosocials' decisions were marked by strong amygdala activity, with the DLPFC signal increasing only in defection. Our findings suggest that proselfs' decisions are controlled by DLPFC-mediated deliberative processes, while prosocials' decisions are initially guided by automatic amygdala processes.
Learning to trust: trust and attachment in early psychosis
DOI:10.1017/S0033291716000015
URL
PMID:26898947
[本文引用: 1]
Distrust and social dysfunction are characteristic in psychosis and may arise from attachment insecurity, which is elevated in the disorder. The relationship between trust and attachment in the early stages of psychosis is unknown, yet could help to understand interpersonal difficulties and disease progression. This study aimed to investigate whether trust is reduced in patients with early psychosis and whether this is accounted for by attachment avoidance and attachment anxiety.
Functional connectivity bias of the orbitofrontal cortex in drug-free patients with major depression
DOI:10.1016/j.biopsych.2009.08.022
URL
PMID:19811772
The orbitofrontal cortex (OFC) plays a crucial role in emotion-processing circuits and should therefore also be included in models of the pathophysiology of major depression. The aim of this study was to compare the functional connectivity of the OFC during emotion processing in patients with major depression and healthy control subjects.
Value, pleasure and choice in the ventral prefrontal cortex
DOI:10.1016/j.tics.2010.12.004
URL
PMID:21216655
[本文引用: 8]
Rapid advances have recently been made in understanding how value-based decision-making processes are implemented in the brain. We integrate neuroeconomic and computational approaches with evidence on the neural correlates of value and experienced pleasure to describe how systems for valuation and decision-making are organized in the prefrontal cortex of humans and other primates. We show that the orbitofrontal and ventromedial prefrontal (VMPFC) cortices compute expected value, reward outcome and experienced pleasure for different stimuli on a common value scale. Attractor networks in VMPFC area 10 then implement categorical decision processes that transform value signals into a choice between the values, thereby guiding action. This synthesis of findings across fields provides a unifying perspective for the study of decision-making processes in the brain.
Neural correlates of social exchanges during the prisoner’s dilemma game in depression
DOI:10.1017/S0033291715002834
URL
PMID:26763141
[本文引用: 6]
Depression is a disabling disorder that significantly impacts on the interpersonal functioning of individuals. However, little is known about the neural substrates of such difficulties. In the last few years neuroeconomics, which combines imaging with multiplayer behavioural economic paradigms, has been used to study the neural substrates of normal and abnormal interpersonal interactions.
Reappraising the ultimatum: An fMRI study of emotion regulation and decision making
DOI:10.1093/cercor/bhs028
URL
PMID:22368088
[本文引用: 2]
Emotion regulation strategies provide a means by which to modulate our social behavior. In this study, we investigated the effect of using reappraisal to both up- and downregulate social decision making. After being instructed on how to use reappraisal, participants played the Ultimatum Game while undergoing functional magnetic resonance imaging and applied the strategies of upregulation (reappraising the proposer's intentions as more negative), down-regulation (reappraising the proposer's intentions as less negative), as well as a baseline &quot;look&quot; condition. As hypothesized, when reappraising, decision acceptance rates were altered, with a greater number of unfair offers accepted while down-regulating and a greater number of unfair offers rejected while upregulating, both relative to the baseline condition. At the neural level, during reappraisal, significant activations were observed in the inferior and middle frontal gyrus (MFG), in addition to the medial prefrontal cortex and cingulate gyrus for unfair offers only. Regulated decisions involved left inferior frontal gyrus for upregulation and MFG for down-regulation strategies, respectively. Importantly, the effects of emotion modulation were evident in posterior insula, with less activation for down-regulation and more activation for upregulation in these areas. Notably, we show for the first time that top-down strategies such as reappraisal strongly affect our socioeconomic decisions.
Mood-congruent cognitive bias in depressed individuals
抑郁症人群的心境一致性认知偏向
Perceived emotional social support in bereaved spouses mediates the relationship between anxiety and depression
DOI:10.1016/j.jad.2017.01.011
URL
PMID:28103522
[本文引用: 1]
Prior research has shown that anxiety symptoms predict later depression symptoms following bereavement. Nevertheless, no research has investigated mechanisms of the temporal relationship between anxiety and later depressive symptoms or examined the impact of depressive symptoms on later anxiety symptoms following bereavement.
Leader emergence through interpersonal neural synchronization
DOI:10.1073/pnas.1422930112
URL
PMID:25831535
[本文引用: 1]
The neural mechanism of leader emergence is not well understood. This study investigated (i) whether interpersonal neural synchronization (INS) plays an important role in leader emergence, and (ii) whether INS and leader emergence are associated with the frequency or the quality of communications. Eleven three-member groups were asked to perform a leaderless group discussion (LGD) task, and their brain activities were recorded via functional near infrared spectroscopy (fNIRS)-based hyperscanning. Video recordings of the discussions were coded for leadership and communication. Results showed that the INS for the leader-follower (LF) pairs was higher than that for the follower-follower (FF) pairs in the left temporo-parietal junction (TPJ), an area important for social mentalizing. Although communication frequency was higher for the LF pairs than for the FF pairs, the frequency of leader-initiated and follower-initiated communication did not differ significantly. Moreover, INS for the LF pairs was significantly higher during leader-initiated communication than during follower-initiated communications. In addition, INS for the LF pairs during leader-initiated communication was significantly correlated with the leaders' communication skills and competence, but not their communication frequency. Finally, leadership could be successfully predicted based on INS as well as communication frequency early during the LGD (before half a minute into the task). In sum, this study found that leader emergence was characterized by high-level neural synchronization between the leader and followers and that the quality, rather than the frequency, of communications was associated with synchronization. These results suggest that leaders emerge because they are able to say the right things at the right time.
Predictors of response to repetitive transcranial magnetic stimulation in depression: A review of recent updates
DOI:10.9758/cpn.2019.17.1.25
URL
PMID:30690937
[本文引用: 1]
Transcranial magnetic stimulation (TMS) has been increasingly used in the treatment of various neuropsychiatric disorders including depression over the past two decades. The responses to treatment with TMS are variable as found in the recent studies. Evidences suggest that various factors influence the outcome of depression treated with TMS. Understanding the predictors of response to TMS treatment in depression will guide the clinician in appropriate selection of patients for TMS treatment as well as needful modification in the TMS technique and protocol to have a better clinical outcome. This article comprehensively reviews the factors that predict the outcome of TMS treatment in depression.
Diminishing reciprocal fairness by disrupting the right prefrontal cortex
DOI:10.1126/science.1129156
URL
PMID:17023614
[本文引用: 2]
Humans restrain self-interest with moral and social values. They are the only species known to exhibit reciprocal fairness, which implies the punishment of other individuals' unfair behaviors, even if it hurts the punisher's economic self-interest. Reciprocal fairness has been demonstrated in the Ultimatum Game, where players often reject their bargaining partner's unfair offers. Despite progress in recent years, however, little is known about how the human brain limits the impact of selfish motives and implements fair behavior. Here we show that disruption of the right, but not the left, dorsolateral prefrontal cortex (DLPFC) by low-frequency repetitive transcranial magnetic stimulation substantially reduces subjects' willingness to reject their partners' intentionally unfair offers, which suggests that subjects are less able to resist the economic temptation to accept these offers. Importantly, however, subjects still judge such offers as very unfair, which indicates that the right DLPFC plays a key role in the implementation of fairness-related behaviors.
The pulvinar nucleus and antidepressant treatment: Dynamic modeling of antidepressant response and remission with ultra-high field functional MRI
DOI:10.1038/s41380-017-0009-x
URL
PMID:29422521
Functional magnetic resonance imaging (fMRI) successfully disentangled neuronal pathophysiology of major depression (MD), but only a few fMRI studies have investigated correlates and predictors of remission. Moreover, most studies have used clinical outcome parameters from two time points, which do not optimally depict differential response times. Therefore, we aimed to detect neuronal correlates of response and remission in an antidepressant treatment study with 7 T fMRI, potentially harnessing advances in detection power and spatial specificity. Moreover, we modeled outcome parameters from multiple study visits during a 12-week antidepressant fMRI study in 26 acute (aMD) patients compared to 36 stable remitted (rMD) patients and 33 healthy control subjects (HC). During an electrical painful stimulation task, significantly higher baseline activity in aMD compared to HC and rMD in the medial thalamic nuclei of the pulvinar was detected (p = 0.004, FWE-corrected), which was reduced by treatment. Moreover, clinical response followed a sigmoid function with a plateau phase in the beginning, a rapid decline and a further plateau at treatment end. By modeling the dynamic speed of response with fMRI-data, perigenual anterior cingulate activity after treatment was significantly associated with antidepressant response (p &lt; 0.001, FWE-corrected). Temporoparietal junction (TPJ) baseline activity significantly predicted non-remission after 2 antidepressant trials (p = 0.005, FWE-corrected). The results underline the importance of the medial thalamus, attention networks in MD and antidepressant treatment. Moreover, by using a sigmoid model, this study provides a novel method to analyze the dynamic nature of response and remission for future trials.
Activation of the human orbitofrontal cortex to a liquid food stimulus is correlated with its subjective pleasantness
DOI:10.1093/cercor/13.10.1064
URL
PMID:12967923
[本文引用: 2]
Single-neuron recording studies in non-human primates indicate that orbitofrontal cortex neurons represent the reward value of the sight, smell and taste of food, and even changes in the relative reward value, but provide no direct evidence on brain activity that is correlated with subjective reports of the pleasantness of food. In this fMRI investigation we report a significant correlation between the activation of a region of the human orbitofrontal cortex and the decrease in subjective pleasantness when a liquid food is eaten to satiety. Moreover, a cluster of voxels in the orbitofrontal cortex showed a decrease in its activation that was specific to the particular liquid food consumed in a meal, providing a neural correlate of sensory-specific satiety to a liquid whole food in humans. This sensory-specific reduction in activation of the orbitofrontal cortex correlating with subjective pleasantness is consistent with an important role for the orbitofrontal cortex in human emotion and motivation, and associated subjective states.
Social functioning in major depressive disorder
DOI:10.1016/j.neubiorev.2016.07.002
URL
PMID:27395342
[本文引用: 2]
Depression is associated with social risk factors, social impairments and poor social functioning. This paper gives an overview of these social aspects using the NIMH Research and Domain Criteria 'Systems for Social Processes' as a framework. In particular, it describes the bio-psycho-social interplay regarding impaired affiliation and attachment (social anhedonia, hyper-sensitivity to social rejection, competition avoidance, increased altruistic punishment), impaired social communication (impaired emotion recognition, diminished cooperativeness), impaired social perception (reduced empathy, theory-of-mind deficits) and their impact on social networks and the use of social media. It describes these dysfunctional social processes at the behavioural, neuroanatomical, neurochemical and genetic levels, and with respect to animal models of social stress. We discuss the diagnostic specificity of these social deficit constructs for depression and in relation to depression severity. Since social factors are importantly involved in the pathogenesis and the consequences of depression, such research will likely contribute to better diagnostic assessments and concepts, treatments and preventative strategies both at the diagnostic and transdiagnostic level.
Automated Talairach atlas labels for functional brain mapping
DOI:10.1002/1097-0193(200007)10:3&lt;120::aid-hbm30&gt;3.0.co;2-8
URL
PMID:10912591
[本文引用: 1]
An automated coordinate-based system to retrieve brain labels from the 1988 Talairach Atlas, called the Talairach Daemon (TD), was previously introduced [Lancaster et al., 1997]. In the present study, the TD system and its 3-D database of labels for the 1988 Talairach atlas were tested for labeling of functional activation foci. TD system labels were compared with author-designated labels of activation coordinates from over 250 published functional brain-mapping studies and with manual atlas-derived labels from an expert group using a subset of these activation coordinates. Automated labeling by the TD system compared well with authors' labels, with a 70% or greater label match averaged over all locations. Author-label matching improved to greater than 90% within a search range of +/-5 mm for most sites. An adaptive grey matter (GM) range-search utility was evaluated using individual activations from the M1 mouth region (30 subjects, 52 sites). It provided an 87% label match to Brodmann area labels (BA 4 &amp; BA 6) within a search range of +/-5 mm. Using the adaptive GM range search, the TD system's overall match with authors' labels (90%) was better than that of the expert group (80%). When used in concert with authors' deeper knowledge of an experiment, the TD system provides consistent and comprehensive labels for brain activation foci. Additional suggested applications of the TD system include interactive labeling, anatomical grouping of activation foci, lesion-deficit analysis, and neuroanatomy education.
Interplay between prior knowledge and communication mode on teaching effectiveness: Interpersonal neural synchronization as a neural marker
DOI:10.1016/j.neuroimage.2019.03.004
URL
PMID:30851445
[本文引用: 2]
Teacher-student interaction allows students to combine prior knowledge with new information to develop new knowledge. It is widely understood that both communication mode and students' knowledge state contribute to the teaching effectiveness (i.e., higher students' scores), but the nature of the interplay of these factors and the underlying neural mechanism remain unknown. In the current study, we manipulated the communication modes (face-to-face [FTF] communication mode/computer-mediated communication [CMC] mode) and prior knowledge states (with vs. without) when teacher-student dyads participated in a teaching task. Using functional near-infrared spectroscopy, the brain activities of both the teacher and student in the dyads were recorded simultaneously. After teaching, perceived teacher-student interaction and teaching effectiveness were assessed. The behavioral results demonstrated that, during teaching with prior knowledge, FTF communication improved students' academic performance, as compared with CMC. Conversely, no such effect was found for teaching without prior knowledge. Accordingly, higher task-related interpersonal neural synchronization (INS) in the left prefrontal cortex (PFC) was found in the FTF teaching condition with prior knowledge. Such INS mediated the relationship between perceived interaction and students' test scores. Furthermore, the cumulative INS in the left PFC could predict the teaching effectiveness early in the teaching process (around 25-35 s into the teaching task) only in FTF teaching with prior knowledge. These findings provide insight into how the interplay between the communication mode and students' knowledge state affects teaching effectiveness. Moreover, our findings suggest that INS could be a possible neuromarker for dynamic evaluation of teacher-student interaction and teaching effectiveness.
Cooperation makes a group be more creative
This study investigated how cooperative and competitive interaction modes affect the group creative performance. The participants were recruited as dyads to solve 2 problems either demanding divergent thinking (alternative uses task, AUT) or not (object characteristic task, OCT). The dyads solved 1 of the 2 problems in the cooperative mode and the other in the competitive mode. Functional near-infrared spectroscopy (fNIRS)-based hyperscanning was used to record their neural activities in the prefrontal and right temporal-parietal junction (r-TPJ) regions. Results revealed the dyads showed higher AUT fluency, AUT originality, OCT fluency, and cooperation level in the cooperative mode than in the competitive mode. The fNIRS data revealed increased (task-baseline) interpersonal brain synchronization (IBS) in the right dorsolateral prefrontal cortex (r-DLPFC) and r-TPJ, only for dyads in the AUT/cooperation condition. In both r-DLPFC and r-TPJ, the IBS of dyads in the AUT/cooperation condition was stronger than in the AUT/competition and OCT/cooperation. Moreover, a stronger IBS was evoked between the regions in prefrontal and posterior temporal regions in the AUT/cooperation condition, as compared with the competition mode. These findings suggest that enhanced IBS may underlie the positive effects of cooperation as compared with the competition in terms of group creativity.
An integrative theory of prefrontal cortex function
DOI:10.1146/annurev.neuro.24.1.167
URL
PMID:11283309
[本文引用: 1]
The prefrontal cortex has long been suspected to play an important role in cognitive control, in the ability to orchestrate thought and action in accordance with internal goals. Its neural basis, however, has remained a mystery. Here, we propose that cognitive control stems from the active maintenance of patterns of activity in the prefrontal cortex that represent goals and the means to achieve them. They provide bias signals to other brain structures whose net effect is to guide the flow of activity along neural pathways that establish the proper mappings between inputs, internal states, and outputs needed to perform a given task. We review neurophysiological, neurobiological, neuroimaging, and computational studies that support this theory and discuss its implications as well as further issues to be addressed
Understanding the minds of others: A neuroimaging meta-analysis
DOI:10.1016/j.neubiorev.2016.03.020
URL
PMID:27073047
[本文引用: 4]
Theory of mind (ToM) is an important skill that refers broadly to the capacity to understand the mental states of others. A large number of neuroimaging studies have focused on identifying the functional brain regions involved in ToM, but many important questions remain with respect to the neural networks implicated in specific types of ToM tasks. In the present study, we conducted a series of activation likelihood estimation (ALE) meta-analyses on 144 datasets (involving 3150 participants) to address these questions. The ALE results revealed common regions shared across all ToM tasks and broader task parameters, but also some important dissociations. In terms of commonalities, consistent activation was identified in the medial prefrontal cortex and bilateral temporoparietal junction. On the other hand, ALE contrast analyses on our dataset, as well as meta-analytic connectivity modelling (MACM) analyses on the BrainMap database, indicated that different types of ToM tasks reliably elicit activity in unique brain areas. Our findings provide the most accurate picture to date of the neural networks that underpin ToM function.
Imbalance in subregional connectivity of the right temporoparietal junction in major depression
DOI:10.1002/hbm.23217
URL
PMID:27090056
[本文引用: 3]
Major depressive disorder (MDD) involves impairment in cognitive and interpersonal functioning. The right temporoparietal junction (RTPJ) is a key brain region subserving cognitive-attentional and social processes. Yet, findings on the involvement of the RTPJ in the pathophysiology of MDD have so far been controversial. Recent connectivity-based parcellation data revealed a topofunctional dualism within the RTPJ, linking its anterior and posterior part (aRTPJ/pRTPJ) to antagonistic brain networks for attentional and social processing, respectively. Comparing functional resting-state connectivity of the aRTPJ and pRTPJ in 72 MDD patients and 76 well-matched healthy controls, we found a seed (aRTPJ/pRTPJ) × diagnosis (MDD/controls) interaction in functional connectivity for eight regions. Employing meta-data from a large-scale neuroimaging database, functional characterization of these regions exhibiting differentially altered connectivity with the aRTPJ/pRTPJ revealed associations with cognitive (dorsolateral prefrontal cortex, parahippocampus) and behavioral (posterior medial frontal cortex) control, visuospatial processing (dorsal visual cortex), reward (subgenual anterior cingulate cortex, medial orbitofrontal cortex, posterior cingulate cortex), as well as memory retrieval and social cognition (precuneus). These findings suggest that an imbalance in connectivity of subregions, rather than disturbed connectivity of the RTPJ as a whole, characterizes the connectional disruption of the RTPJ in MDD. This imbalance may account for key symptoms of MDD in cognitive, emotional, and social domains. Hum Brain Mapp 37:2931-2942, 2016. © 2016 Wiley Periodicals, Inc.
Social-economical decision making in current and remitted major depression
DOI:10.1017/S0033291714002414
URL
PMID:25300570
Prosocial emotions related to self-blame are important in guiding human altruistic decisions. These emotions are elevated in major depressive disorder (MDD), such that MDD has been associated with guilt-driven pathological hyper-altruism. However, the impact of such emotional impairments in MDD on different types of social decision-making is unknown.
Brain-to-brain synchrony in parent-child dyads and the relationship with emotion regulation revealed by fNIRS-based hyperscanning
DOI:10.1016/j.neuroimage.2018.05.060
URL
PMID:29807152
[本文引用: 2]
Parent-child synchrony, the coupling of behavioral and biological signals during social contact, may fine-tune the child's brain circuitries associated with emotional bond formation and the child's development of emotion regulation. Here, we examined the neurobiological underpinnings of these processes by measuring parent's and child's prefrontal neural activity concurrently with functional near-infrared spectroscopy hyperscanning. Each child played both a cooperative and a competitive game with the parent, mostly the mother, as well as an adult stranger. During cooperation, parent's and child's brain activities synchronized in the dorsolateral prefrontal and frontopolar cortex (FPC), which was predictive for their cooperative performance in subsequent trials. No significant brain-to-brain synchrony was observed in the conditions parent-child competition, stranger-child cooperation and stranger-child competition. Furthermore, parent-child compared to stranger-child brain-to-brain synchrony during cooperation in the FPC mediated the association between the parent's and the child's emotion regulation, as assessed by questionnaires. Thus, we conclude that brain-to-brain synchrony may represent an underlying neural mechanism of the emotional connection between parent and child, which is linked to the child's development of adaptive emotion regulation. Future studies may uncover whether brain-to-brain synchrony can serve as a neurobiological marker of the dyad's socio-emotional interaction, which is sensitive to risk conditions, and can be modified by interventions.
Predictive value of dorso-lateral prefrontal connectivity for rTMS response in treatment-resistant depression: A brain perfusion SPECT study
DOI:10.1016/j.brs.2018.05.010
URL
PMID:29802071
[本文引用: 3]
Previous clinical trials have suggested that repetitive transcranial magnetic stimulation (rTMS) has a significant antidepressant effect in patients with treatment resistant depression (TRD). However, results remain heterogeneous with many patients without effective response.
A neural basis for social cooperation
DOI:10.1016/s0896-6273(02)00755-9
URL
PMID:12160756
Cooperation based on reciprocal altruism has evolved in only a small number of species, yet it constitutes the core behavioral principle of human social life. The iterated Prisoner's Dilemma Game has been used to model this form of cooperation. We used fMRI to scan 36 women as they played an iterated Prisoner's Dilemma Game with another woman to investigate the neurobiological basis of cooperative social behavior. Mutual cooperation was associated with consistent activation in brain areas that have been linked with reward processing: nucleus accumbens, the caudate nucleus, ventromedial frontal/orbitofrontal cortex, and rostral anterior cingulate cortex. We propose that activation of this neural network positively reinforces reciprocal altruism, thereby motivating subjects to resist the temptation to selfishly accept but not reciprocate favors.
Neural correlates of social cooperation and non-cooperation as a function of psychopathy
DOI:10.1016/j.biopsych.2006.07.021
URL
PMID:17046722
[本文引用: 1]
Psychopathy is a disorder involving a failure to experience many emotions that are necessary for appropriate social behavior. In this study, we probed the behavioral, emotional, and neural correlates of psychopathic traits within the context of a dyadic social interaction.
The neuroscience of social decision-making
DOI:10.1146/annurev.psych.121208.131647
URL
PMID:20822437
[本文引用: 2]
Given that we live in highly complex social environments, many of our most important decisions are made in the context of social interactions. Simple but sophisticated tasks from a branch of experimental economics known as game theory have been used to study social decision-making in the laboratory setting, and a variety of neuroscience methods have been used to probe the underlying neural systems. This approach is informing our knowledge of the neural mechanisms that support decisions about trust, reciprocity, altruism, fairness, revenge, social punishment, social norm conformity, social learning, and competition. Neural systems involved in reward and reinforcement, pain and punishment, mentalizing, delaying gratification, and emotion regulation are commonly recruited for social decisions. This review also highlights the role of the prefrontal cortex in prudent social decision-making, at least when social environments are relatively stable. In addition, recent progress has been made in understanding the neural bases of individual variation in social decision-making.
Neural mechanisms of reinforcement learning in unmedicated patients with major depressive disorder
DOI:10.1093/brain/awx025
URL
PMID:28334960
[本文引用: 1]
According to current concepts, major depressive disorder is strongly related to dysfunctional neural processing of motivational information, entailing impairments in reinforcement learning. While computational modelling can reveal the precise nature of neural learning signals, it has not been used to study learning-related neural dysfunctions in unmedicated patients with major depressive disorder so far. We thus aimed at comparing the neural coding of reward and punishment prediction errors, representing indicators of neural learning-related processes, between unmedicated patients with major depressive disorder and healthy participants. To this end, a group of unmedicated patients with major depressive disorder (n = 28) and a group of age- and sex-matched healthy control participants (n = 30) completed an instrumental learning task involving monetary gains and losses during functional magnetic resonance imaging. The two groups did not differ in their learning performance. Patients and control participants showed the same level of prediction error-related activity in the ventral striatum and the anterior insula. In contrast, neural coding of reward prediction errors in the medial orbitofrontal cortex was reduced in patients. Moreover, neural reward prediction error signals in the medial orbitofrontal cortex and ventral striatum showed negative correlations with anhedonia severity. Using a standard instrumental learning paradigm we found no evidence for an overall impairment of reinforcement learning in medication-free patients with major depressive disorder. Importantly, however, the attenuated neural coding of reward in the medial orbitofrontal cortex and the relation between anhedonia and reduced reward prediction error-signalling in the medial orbitofrontal cortex and ventral striatum likely reflect an impairment in experiencing pleasure from rewarding events as a key mechanism of anhedonia in major depressive disorder.
The neurobiology of rewards and values in social decision making
DOI:10.1038/nrn3776
URL
PMID:24986556
[本文引用: 2]
How does our brain choose the best course of action? Choices between material goods are thought to be steered by neural value signals that encode the rewarding properties of the choice options. Social decisions, by contrast, are traditionally thought to rely on neural representations of the self and others. However, recent studies show that many types of social decisions may also involve neural value computations. This suggests a unified mechanism for motivational control of behaviour that may incorporate both social and non-social factors. In this Review, we outline a theoretical framework that may help to identify possible overlaps and differences between the neural processes that guide social and non-social decision making.
Effects of early life stress on depression, cognitive performance and brain morphology
DOI:10.1017/S0033291716002403
URL
PMID:27682320
[本文引用: 1]
Childhood early life stress (ELS) increases risk of adulthood major depressive disorder (MDD) and is associated with altered brain structure and function. It is unclear whether specific ELSs affect depression risk, cognitive function and brain structure.
Dimensional depression severity in women with major depression and post-traumatic stress disorder correlates with fronto-amygdalar hypoconnectivty
DOI:10.1038/mp.2015.149
URL
PMID:26416545
[本文引用: 1]
Depressive symptoms are common in multiple psychiatric disorders and are frequent sequelae of trauma. A dimensional conceptualization of depression suggests that symptoms should be associated with a continuum of deficits in specific neural circuits. However, most prior investigations of abnormalities in functional connectivity have typically focused on a single diagnostic category using hypothesis-driven seed-based analyses. Here, using a sample of 105 adult female participants from three diagnostic groups (healthy controls, n=17; major depression, n=38; and post-traumatic stress disorder, n=50), we examine the dimensional relationship between resting-state functional dysconnectivity and severity of depressive symptoms across diagnostic categories using a data-driven analysis (multivariate distance-based matrix regression). This connectome-wide analysis identified foci of dysconnectivity associated with depression severity in the bilateral amygdala. Follow-up seed analyses using subject-specific amygdala segmentations revealed that depression severity was associated with amygdalo-frontal hypo-connectivity in a network of regions including bilateral dorsolateral prefrontal cortex, anterior cingulate and anterior insula. In contrast, anxiety was associated with elevated connectivity between the amygdala and the ventromedial prefrontal cortex. Taken together, these results emphasize the centrality of the amygdala in the pathophysiology of depressive symptoms, and suggest that dissociable patterns of amygdalo-frontal dysconnectivity are a critical neurobiological feature across clinical diagnostic categories.
Construction of a 3D probabilistic atlas of human cortical structures
DOI:10.1016/j.neuroimage.2007.09.031
URL
PMID:18037310
[本文引用: 1]
We describe the construction of a digital brain atlas composed of data from manually delineated MRI data. A total of 56 structures were labeled in MRI of 40 healthy, normal volunteers. This labeling was performed according to a set of protocols developed for this project. Pairs of raters were assigned to each structure and trained on the protocol for that structure. Each rater pair was tested for concordance on 6 of the 40 brains; once they had achieved reliability standards, they divided the task of delineating the remaining 34 brains. The data were then spatially normalized to well-known templates using 3 popular algorithms: AIR5.2.5's nonlinear warp (Woods et al., 1998) paired with the ICBM452 Warp 5 atlas (Rex et al., 2003), FSL's FLIRT (Smith et al., 2004) was paired with its own template, a skull-stripped version of the ICBM152 T1 average; and SPM5's unified segmentation method (Ashburner and Friston, 2005) was paired with its canonical brain, the whole head ICBM152 T1 average. We thus produced 3 variants of our atlas, where each was constructed from 40 representative samples of a data processing stream that one might use for analysis. For each normalization algorithm, the individual structure delineations were then resampled according to the computed transformations. We next computed averages at each voxel location to estimate the probability of that voxel belonging to each of the 56 structures. Each version of the atlas contains, for every voxel, probability densities for each region, thus providing a resource for automated probabilistic labeling of external data types registered into standard spaces; we also computed average intensity images and tissue density maps based on the three methods and target spaces. These atlases will serve as a resource for diverse applications including meta-analysis of functional and structural imaging data and other bioinformatics applications where display of arbitrary labels in probabilistically defined anatomic space will facilitate both knowledge-based development and visualization of findings from multiple disciplines.
Accelerated TMS for depression: A systematic review and meta-analysis
DOI:10.1016/j.psychres.2018.12.041
URL
PMID:31207865
Repetitive transcranial magnetic stimulation (TMS) is now widely available for the clinical treatment of depression, but the associated financial and time burdens are problematic for patients. Accelerated TMS (aTMS) protocols address these burdens and attempt to increase the efficiency of standard TMS. This systematic review and meta-analysis aimed to examine accelerated TMS studies for depressive disorders in accordance with PRISMA guidelines. Inclusion criteria consisted of studies with full text publications available in English describing more than one session of TMS (repetitive or theta burst stimulation) per day. Studies describing accelerated TMS protocols for conditions other than depression or alternative neuromodulation methods, preclinical studies, and neurophysiology studies regarding transcranial stimulation were excluded. Eighteen articles describing eleven distinct studies (seven publications described overlapping samples) met eligibility criteria. A Hedges' g effect size and confidence intervals were calculated. The summary analysis of three suitable randomized control trials revealed a cumulative effect size of 0.39 (95% CI 0.005-0.779). A separate analysis including open-label trials and active arms of suitable RCTs revealed a g of 1.27 (95% CI 0.902-1.637). Overall, the meta-analysis suggested that aTMS improves depressive symptom severity. In general, study methodologies were acceptable, but future efforts could enhance sham techniques and blinding.
The influence of cooperation and defection on social decision making in depression: A study of the iterated prisoner's dilemma game
DOI:10.1016/j.psychres.2016.10.025
URL
PMID:27821362
[本文引用: 3]
This study evaluated the influence of self-reported levels of depression on interpersonal strategic decision making when interacting with partners who differed in their predetermined tendency to cooperate in three separate computerized iterated Prisoner's Dilemma Games (iPDGs). Across 29 participants, cooperation was lowest when interacting with a predominantly defecting partner and highest when interacting with a predominantly cooperating partner. Greater depression severity was related to steadier and continued cooperation over trials with the cooperating partner, seeming to reflect a prosocial response tendency when interacting with this partner. With the unbiased partner, depression severity was associated with a more volatile response pattern in reaction to cooperation and defection by this partner. Severity of depression did not influence cooperation with a defecting partner or expectations about partner cooperation reported before the task began. Taken together, these data appear to show that in predominately positive interactions, as in the cooperating partner condition, depression is associated with less volatile, more consistent cooperation. When such clear feedback is absent, as in the unbiased partner condition, depression is associated with more volatile behavior. Nonetheless, participants were generally able to adapt their behavior accordingly in this dynamic interpersonal decision making context.
The importance of the lateral prefrontal cortex for strategic decision making in the prisoner’s dilemma
Attenuated intrinsic connectivity within cognitive control network among individuals with remitted depression: Temporal stability and association with negative cognitive styles
DOI:10.1002/hbm.23564
URL
PMID:28345197
Many individuals with major depressive disorder (MDD) experience cognitive dysfunction including impaired cognitive control and negative cognitive styles. Functional connectivity magnetic resonance imaging studies of individuals with current MDD have documented altered resting-state connectivity within the default-mode network and across networks. However, no studies to date have evaluated the extent to which impaired connectivity within the cognitive control network (CCN) may be present in remitted MDD (rMDD), nor have studies examined the temporal stability of such attenuation over time. This represents a major gap in understanding stable, trait-like depression risk phenotypes. In this study, resting-state functional connectivity data were collected from 52 unmedicated young adults with rMDD and 47 demographically matched healthy controls, using three bilateral seeds in the CCN (dorsolateral prefrontal cortex, inferior parietal lobule, and dorsal anterior cingulate cortex). Mean connectivity within the entire CCN was attenuated among individuals with rMDD, was stable and reliable over time, and was most pronounced with the right dorsolateral prefrontal cortex and right inferior parietal lobule, results that were corroborated by supplemental independent component analysis. Attenuated connectivity in rMDD appeared to be specific to the CCN as opposed to representing attenuated within-network coherence in other networks (e.g., default-mode, salience). In addition, attenuated connectivity within the CCN mediated relationships between rMDD status and cognitive risk factors for depression, including ruminative brooding, pessimistic attributional style, and negative automatic thoughts. Given that these cognitive markers are known predictors of relapse, these results suggest that attenuated connectivity within the CCN could represent a biomarker for trait phenotypes of depression risk. Hum Brain Mapp 38:2939-2954, 2017. © 2017 Wiley Periodicals, Inc.
Be nice if you have to—The neurobiological roots of strategic fairness
DOI:10.1093/scan/nsu114
URL
PMID:25190704
[本文引用: 2]
Social norms, such as treating others fairly regardless of kin relations, are essential for the functioning of human societies. Their existence may explain why humans, among all species, show unique patterns of prosocial behaviour. The maintenance of social norms often depends on external enforcement, as in the absence of credible sanctioning mechanisms prosocial behaviour deteriorates quickly. This sanction-dependent prosocial behaviour suggests that humans strategically adapt their behaviour and act selfishly if possible but control selfish impulses if necessary. Recent studies point at the role of the dorsolateral prefrontal cortex (DLPFC) in controlling selfish impulses. We test whether the DLPFC is indeed involved in the control of selfish impulses as well as the strategic acquisition of this control mechanism. Using repetitive transcranial magnetic stimulation, we provide evidence for the causal role of the right DLPFC in strategic fairness. Because the DLPFC is phylogenetically one of the latest developed neocortical regions, this could explain why complex norm systems exist in humans but not in other social animals.
The neural responses to social cooperation in gain and loss context
DOI:10.1371/journal.pone.0160503
URL
PMID:27494142
[本文引用: 2]
Cooperation is pervasive and constitutes the core behavioral principle of human social life. Previous studies have revealed that mutual cooperation was reliably correlated with two reward-related brain regions, the ventral striatum and the orbitofrontal cortex. Using functional magnetic resonance imaging (fMRI), this study sought to investigate how the loss and gain contexts modulated the neural responses to mutual cooperation. Twenty-five female participants were scanned when they played a series of one-shot prisoner's dilemma games in the loss and gain contexts. Specifically, participants and partners independently chose to either cooperate with each other or not, and each was awarded or deprived of (in the gain context or the loss context, respectively) a sum of money which depended upon the interaction of their choices. Behavioral results indicated that participants cooperated in nearly half of the experiment trials and reported higher level of positive emotions for mutual cooperation in both contexts, but they cooperated more in the gain than in the loss context. At the neural level, stronger activities in the orbitofrontal cortex were observed for mutual cooperation compared with the other three outcomes in both contexts, while stronger activation in ventral striatum associated with mutual cooperation was observed in the gain context only. Together, our data indicated that, even in the one-shot interaction under loss context, participants still exhibited preference for cooperation and the rewarding experience from a mutually cooperative social interaction activated the ventral striatum and the orbitofrontal cortex, but the loss context weakened the association between the ventral striatum activation and mutual cooperation.
Individual differences in rumination in healthy and depressive samples: association with brain structure, functional connectivity and depression
DOI:10.1017/S0033291715000938
URL
PMID:26219340
[本文引用: 2]
Rumination is an important cognitive risk factor for onset and relapse of depression. However, no studies have employed a dimensional approach in investigating the neural correlates of rumination and the relationship with depression.
Common and distinct patterns of grey-matter volume alteration in major depression and bipolar disorder: evidence from voxel-based meta-analysis
DOI:10.1038/mp.2016.72
URL
PMID:27217146
Finding robust brain substrates of mood disorders is an important target for research. The degree to which major depression (MDD) and bipolar disorder (BD) are associated with common and/or distinct patterns of volumetric changes is nevertheless unclear. Furthermore, the extant literature is heterogeneous with respect to the nature of these changes. We report a meta-analysis of voxel-based morphometry (VBM) studies in MDD and BD. We identified studies published up to January 2015 that compared grey matter in MDD (50 data sets including 4101 individuals) and BD (36 data sets including 2407 individuals) using whole-brain VBM. We used statistical maps from the studies included where available and reported peak coordinates otherwise. Group comparisons and conjunction analyses identified regions in which the disorders showed common and distinct patterns of volumetric alteration. Both disorders were associated with lower grey-matter volume relative to healthy individuals in a number of areas. Conjunction analysis showed smaller volumes in both disorders in clusters in the dorsomedial and ventromedial prefrontal cortex, including the anterior cingulate cortex and bilateral insula. Group comparisons indicated that findings of smaller grey-matter volumes relative to controls in the right dorsolateral prefrontal cortex and left hippocampus, along with cerebellar, temporal and parietal regions were more substantial in major depression. These results suggest that MDD and BD are characterised by both common and distinct patterns of grey-matter volume changes. This combination of differences and similarities has the potential to inform the development of diagnostic biomarkers for these conditions.
Cooperation makes two less-creative individuals turn into a highly-creative pair
DOI:10.1016/j.neuroimage.2018.02.007
URL
PMID:29427846
[本文引用: 1]
This study aimed to investigate which type of group (e.g., consisting of less-creative or highly-creative individuals) would perform better in solving creativity problems, and explore the underlying inter-brain neural correlates between team members. A preliminary test (an alternative-uses task) was performed to rank individuals' level of creativity, and divide participants into three types of dyads: high-high (two highly-creative individuals), low-low (two less-creative individuals), and high-low (one highly-creative and one less-creative individual). Dyads were then asked to solve a realistic presented problem (RPP; a typical creativity problem) during which a functional near-infrared spectroscopy (fNIRS)-based hyperscanning device was used to record the variation of interpersonal brain synchronization (IBS). Results revealed that less-creative individuals, while working together, would perform as well as highly-creative individuals. The low-low dyads showed higher levels of cooperation behaviour than the other two types of dyads. The fNIRS results revealed increased IBS only for low-low dyads at PFC (prefrontal cortex) and rTPJ (right temporal-parietal junction) brain regions during RPP task performance. In the rDLPFC (right dorsolateral prefrontal cortex), the IBS in the low-low dyads was stronger than that of high-high and high-low dyads. In the rTPJ, the IBS in the low-low dyads was only stronger than that of the high-low dyads. Besides, the IBS at rDLPFC and rTPJ regions in the low-low dyads was positively correlated with their cooperation behaviour and group creative performance. These findings indicated when two less-creative individuals worked on a creativity problem together, they tended to cooperate with each other (indicated by both behaviour index and increased IBS at rDLPFC and rTPJ), which benefited their creative performance.
The dynamics of belief updating in human cooperation: Findings from inter-brain ERP hyperscanning
DOI:10.1016/j.neuroimage.2019.05.029
URL
PMID:31085300
[本文引用: 1]
Generally, successful cooperation can only be established when the interacting persons believe that they would not be betrayed; this belief can be updated by observing the other persons' actual choices. Thus, the process of belief updating plays an important role in conditional cooperation. Using the Prisoner's Dilemma Game (PDG) with event-related potential (ERP) hyperscanning, this study investigated the dynamics of belief updating in a dyad. During the task, participants were asked if they believed that their opponent would cooperate in the next trial, and their answers functioned as a self-reported index of reciprocal belief. The results suggested that this index shows strong associations with participants' behavioral choices (cooperate/betray). At the individual level, the amplitudes of the ERP components frontal P3a and parietal P3b elicited by the decision outcome were sensitive to belief updating. At the interpersonal level, the between-subject synchronization in P3b was higher than those in the other conditions when the paired participants confirmed each other's reciprocal beliefs. Since previous studies have linked the P3b with memory updating, we suggest that a cooperative relationship is built up when the memory systems (which support belief updating) of two interacting persons reach a high level of coordination. These findings may help explain how conditional cooperation develops between strangers.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}