

1 前言
程序性运动学习是指通过重复执行运动任务, 借助骨骼、肌肉以及相应的神经反射获得新知识的过程。根据学习内容呈现顺序的不同可将其分为序列学习和随机学习两种形式, 其中序列学习通过一系列不断重复的、固定长度的序列循环呈现而发生学习(杨光, 2014), 随机学习则是在无序的呈现中实现学习的过程(Pascual-Leone, Wassermann, Grafman, & Hallett, 1996)。程序性运动学习不仅是人类日常活动的重要组成部分, 更是运动技能习得的重要方式(Clegg, Digirolamo, & Keele, 1998)。它为我们探究复杂形式下的运动学习提供了范式, 能够反映人类大脑运动认知系统在不同状态下的适应性与可塑性(Grafton, Woods, & Tyszka, 1994)。
研究表明, 程序性运动学习涉及多个脑功能区的协同作用, 包括初级运动皮层(primary motor cortex, M1)、辅助运动区(supplementary motor area, SMA)、前运动区(premotor cortex)和背外侧前额叶(dorsolateral prefrontal cortex, DLPFC)等重要脑功能区(Leonora, Teo, Ignacio, Rothwell, & Marjan, 2010; Poldrack et al., 2005; Schendan, Searl, Melrose, & Stern, 2003; Seidler et al., 2005)。其中, 以M1为代表的运动功能区与以DLPFC为代表的认知功能区之间的联系与投射是解释不同运动学习表现的重要依据(Cao et al., 2018; Lam et al., 2015)。
与右侧前额叶皮质相比, 左侧背外侧前额叶, 尤其是BA46区(Kielan, Peter, & Krakauer, 2009), 被认为是具有整合运动学习和运动控制功能的重要脑区(Miller, 2000)。尽管DLPFC脑功能区参与和运动学习有关的运动认知过程, 但是它以怎样的方式参与运动认知的问题仍然未能得到回答。神经影像学研究显示, 在序列学习任务中DLPFC出现持续性的激活(Jenkins, Brooks, Nixon, Frackowiak, & Passingham, 1994; Sakai et al., 1998; Toni, Krams, Turner, & Passingham, 1998)。Shadmehr和Holcomb (1997)报告在序列和随机两种学习任务中, 序列学习表现提升的同时前额叶与运动皮层都被激活。根据Hikosaka理论, DLPFC参与空间序列获取的过程, 即前额叶皮层涉及处理最初的感觉输入以及描绘空间序列的过程, 最终到达运动皮层(Hikosaka, Nakamura, Sakai, & Nakahara, 2002)。反之, 如果测试中未包含序列学习任务, 前额叶皮层没有发生激活(Willingham, 1998)。这些结果都表明了DLPFC的激活因任务情境而异。当采用非侵入性脑刺激破坏DLPFC脑区兴奋性后, 被试在空间信息处理任务中按键反应时间增加, 学习效果被破坏(Robertson, Tormos, Maeda, & Pascual-Leone, 2001), 反映了DLPFC兴奋性对最终运动学习效果的影响。在病理学方面的研究中, 前人发现小脑功能障碍患者、帕金森病患者的程序性学习功能可能受损(Pascual-Leone et al., 1996); 另一方面, 即使颞叶和背内侧丘脑损伤(Gordon, 1988; Squire, 1992)、或者患有阿尔茨海默病的病人(Grafman et al., 1990), 他们的程序性学习功能依旧得以保留。这些结果说明了当与额叶, 特别是与背外侧前额叶相连的皮质受损的情况下, 病人的程序性学习功能受到影响, 表明程序性学习依赖于与背外侧前额叶相连通路的完好性(Fuster & Alexander, 1971)。
已有的研究主要关注DLPFC这一单个脑区在相关任务中的激活状态, 而未从通路角度探讨DLPFC作为认知脑功能区重要结点与作为运动输出最终窗口的M1之间的联通性。尽管背外侧前额叶在运动学习中起到了重要作用, 但是神经影像学研究却无法揭示DLPFC-M1联通性在不同运动学习中的改变及作用(Friston, 2011)。有效的神经功能联结是快速处理信息的前提, 从DLPFC与M1联通性的角度探索不同程序性运动学习是认识运动学习发生规律的重要途径之一。
本研究使用两重位置(Dual-site)两连发经颅磁刺激法(Transcranial magnetic stimulation, TMS), 这一电生理手段能够直接体现一个神经系统对另一个系统施加的有效连接或直接影响从而精确描述可塑性变化(Lafleur, Tremblay, Whittingstall, & Lepage, 2016)。两连发刺激中第一发刺激为条件刺激(Conditioning stimulation, CS), 用于刺激DLPFC区域; 第二发刺激为测试刺激(Test stimulation, TS), 用于刺激M1。通过改变两发刺激之间的时间间隔和测试强度, 能够探测多目标脑区之间神经传导通路的兴奋性, 尤其是获得M1以外特定脑功能区神经回路的性质(Ni & Chen, 2012; Ni, Florian, Chen, & Ziemann, 2011; Ni et al., 2009)。该方法的优势在于能够将不同脑区之间联系起来, 探究它们之间的功能连接性及兴奋或抑制环路, 通过TS诱发的运动诱发电位(Motor evoked potential, MEP)振幅的变化反映在皮质水平上相连接的CS刺激区域到TS刺激区域的功能性通路, 即脑区之间的联通性(Lazzaro et al., 1999; Rothwell, 2011)。随着两连发TMS技术的发展, 该方法目前广泛用于评估非同源区域之间的半球间连接、半球内连接以及小脑皮质连接的科学研究(Ziemann et al., 2015), 以及脑卒中、运动障碍、肌萎缩侧索硬化症等临床诊断中(Ridding & Rothwell, 2007), 具有较高的信效度(Marco, Carlo, & Elena, 2011)。
本研究目的是探讨左侧DLPFC到M1的联通性在两种不同程序性运动学习中的差异。程序性运动学习采用连续反应时间任务(Serial response time task, SRTT) (Leonora et al., 2010), 在这类任务的每个试次中, 目标刺激会出现在四个可能的位置之一, 受试者必须通过按下键盘上的相应键, 对刺激出现位置尽可能快地做出反应。目标刺激出现位置的顺序根据实验目的的不同可以呈现序列或随机的形式, 相比之下, 其他不能将刺激设置成序列的任务例如虚拟机械操纵任务(Krebs, Hogan, Hening, Adamovich, & Poizner, 2001)、追踪转翼任务(Noguchi, Demura, Nagasawa, & Uchiyama, 2009)等则不具备此项优势。因此“顺序可操作性” (Willingham, Salidis, & Gabrieli, 2002)使得其近年来被更广泛地应用于程序性学习研究中, 是比较序列顺序和随机顺序条件下学习的重要行为范式(Mayor-Dubois, Zesiger, van der Linden, & Roulet-Perez, 2016)。实验1通过改变两发刺激的时间间隔探究DLPFC到M1投射的时间过程; 实验2将通过SRTT和TMS的结合, 探索不同运动学习类型对DLPFC-M1输入-输出曲线的影响。
根据前人文献及理论(Hikosaka et al., 2002; Robertson et al., 2001; Toni et al., 1998; Willingham, 1998), DLPFC参与空间序列获取过程并在序列学习中持续激活, 而在无意识的学习中未发生激活, 且抑制DLPFC能够降低按键学习任务的正确率及增加反应时间, 因此本研究提出假设:序列学习导致DLPFC-M1的联通性增强, 且与学习成绩提升相关; 而随机学习没有改变。
2 方法
2.1 被试
44名被试参加了本研究一项或多项实验。21名被试(7名女性)参与了实验1, 平均年龄21.10 ± 1.97岁; 40名被试(15名女性)参与了实验2, 平均年龄21.75 ± 1.74岁。两项实验间隔2周以上保证无实验后效影响(Ni, Gunraj, Kailey, Cash, & Chen, 2014)。所有被试均为右利手(Oldfield, 1971), 视力或者矫正视力正常。所有被试根据赫尔辛基宣言签署了书面知情同意书。TMS实验由当地伦理委员会审核批准。
2.2 实验设计
2.2.1 实验1:左侧DLPFC到M1投射的时间过程
本实验采用单因素重复测量实验设计, 自变量为两重位置两连发TMS中CS和TS的时间间隔, 因变量为两连发MEP振幅。通过对DLPFC和M1连续的两连发刺激, 探究皮质脊髓输出兴奋性的改变。
首先, 实验确定了CS强度和TS强度。根据前人文献, TS (M1)的强度设置为用相同的TMS线圈在静息时诱发振幅为1 mV的MEP时机器输出强度, 以此定为振幅的基本单位, 作为后续两连发TMS诱发MEP振幅的比较对象(Hallett, 2000)。CS (DLPFC)的强度为110%静息运动阈值(Rest motor threshold, RMT)。由已有研究可知, 阈上刺激更能促进额叶与M1之间的相互传导, 而较高的阈上刺激强度则会带来不适感(Jacinta, Catherine, Boorman, Heidi, & Rushworth, 2010; Koch et al., 2007)。正式实验前首先确定每位被试右手第一背侧骨间肌的RMT。RMT指目标肌肉在静息状态下, 连续10次刺激中有至少5次诱发出大于50 μV振幅MEP的最小磁刺激强度。
根据不同脑功能区向M1投射的神经纤维长度(Oh et al., 2014; Wedeen et al., 2012)以及神经递质(Ziemann, 2004)的不同选择10个连续时间间隔(Interstimulus interval, ISI) (2、4、6、8、10、12、15、20、25、30 ms), 由此可以确定在某一特定CS强度下被刺激脑功能区向M1投射的时间过程, 从而探知由此形成的神经回路的性质(兴奋性或抑制性)以及达到兴奋或抑制峰值效应所需时间间隔(图1A, B)。每位被试在每个两连发TMS时间间隔下各进行10个试次, 试次以5秒钟的间歇时间进行, 10个时间间隔以随机顺序呈现, 共100个试次。根据前人文献报告, 试次与试次之间的间隔为5秒钟时, 试次之间不会产生相互影响(Hallett, 2000)。
图1
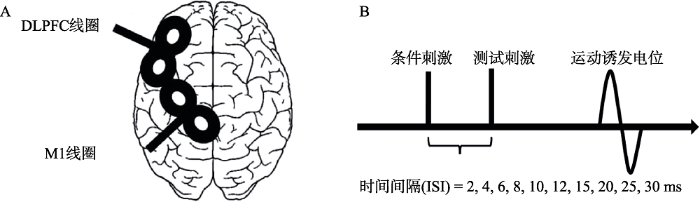
图1
(A)左半球线圈位置示意图(Hasan et al., 2013); (B)两连发TMS示意图
2.2.2 实验2:不同运动学习类型对DLPFC-M1联通性输入-输出曲线(Input-output curve, IO曲线)的影响
实验2中以学习种类(序列学习或随机学习)作为组间因素, 检测在两个最佳的时间间隔下(10和25 ms, 由实验1结果所得), 不同类型的运动学习对DLPFC-M1联通性IO曲线的影响(图2A)。对于学习任务, 以组块为组内变量, 采用2(组别:序列学习组, 随机学习组) × 2(组块:组块1, 组块10)混合设计, 因变量为被试的组块平均反应时; 对生理学指标, 以刺激强度、学习前后测作为组内因素, 采用2(组别:序列学习组, 随机学习组) × 2(前后测:前测, 后测) × 5(刺激强度:单发TMS和双发TMS强度各异)的混合设计, 因变量为MEP振幅。
图2

其中, 刺激强度水平中两连发TMS的CS强度为50% RMT, 70% RMT, 90% RMT, 110% RMT, 130% RMT (CS刺激于DLPFC, 对于阈值上下的强度敏感性高, 所以以RMT为基准) (Lam et al., 2015); 单发TMS的强度设定为70%、90%、100%、110%、120% 1 mV (需要与单发MEP的1 mV标准值来比较, 因此以1 mV的强度为基准), 从而完成IO曲线的测量。每位被试在各个强度水平下均进行10个试次刺激, 刺激强度以每5秒的频率随机出现。实验将以前测-学习任务-后测的顺序进行(图2A), 在学习任务完成后, 序列学习组的被试将被询问是否发现序列的存在, 并复述序列。
2.3 测试方法
2.3.1 两重位置两连发经颅磁刺激
研究选用两个直径为5 cm的小8字线圈, 分别连接两个单脉冲单相TMS刺激器(型号Magstim200, Whitland, Dyfeld, UK), 对左侧DLPFC和左侧M1进行经颅磁刺激。本研究中, 刺激点DLPFC和M1均位于左侧半脑, 从而减少来自半球间的测量偏差, 更专注于优势半球。为了研究左半球DLPFC-M1的联通性, 实验中的CS将对左侧DLPFC施加经颅磁刺激, 通过测量刺激DLPFC后同侧M1输出兴奋性的程度从而量化DLPFC对M1的兴奋性或抑制性的影响。
施加在左侧DLPFC的刺激为CS, 施加在M1的刺激为TS。TS线圈位置为右手第一背侧骨间肌对应在M1区的最佳激活位置, 即“运动热点”。TS线圈的手柄几乎垂直于中央沟, 并与大脑中央矢状线呈30°~45°, 通过大脑的电流为“后-前方向”。为确定左侧背外侧前额叶的精确位置, 与经颅磁刺激仪器连接的脑区导航仪(型号eemagine visor2)通过对被试的大脑核磁共振成像扫描结果进行建模, 依据大脑左侧BA 46脑区的Talairach坐标((x, y, z):-40, 28, 30)确定具体的CS位置(Rowe, Stephan, Friston, Frackowiak, & Passingham, 2005)。核磁共振成像扫描采用西门子3.0 T全身磁共振成像仪(Siemens Magnetom Trio 3.0T)进行结构像数据的采集。采用12导阵列线圈, 以平面回波成像(EPI)序列采集T1*结构像, 扫描参数:TR 2000 ms, TE 30 ms, 断层间隔1 mm, 翻转角度90°, 层厚3 mm, 矩阵64×64, 视野240 mm × 240 mm, 体素大小为3.75 mm × 3.75 mm × 5.0 mm, 共33 层, 隔层扫描(张兰兰 等, 2017)。
2.3.2 肌电图记录
直径9 mm的碗状Ag-AgCl表面电极片用于记录被试右手第一背侧骨间肌表面肌电图。实验时活动电极放置在肌腹, 参考电极放在掌指关节处。数据通过神经放电信号采集器(型号CED Micor1401)采集, 采集到的信号使用电生理信号调节放大器(型号Model 2024F)进行过滤(带通滤波20~ 25000 Hz), 并放大1000倍; 过滤放大后的信号以1 kHz的频率被数字化记录, 通过Signal 6.0软件进行线下分析。
2.3.3 行为实验范式
本实验的行为实验任务采用的是序列反应时任务(SRTT)。使用E-prime软件制作实验程序, 通过笔记本电脑(惠普Pavilion)屏幕(15.6寸, 分辨率1980×1020, 刷新率75 Hz)呈现刺激, 被试使用标准的键盘进行按键反应。被试坐在安静的实验室中, 双眼与屏幕距离约80 cm, 白色的屏幕中间显示一个黑色箭头。箭头可能出现在四个不同的方向(9、11、1和3点钟方向), 每个方向需要被试用不同手指按下对应按钮(分别是:食指、中指、无名指、小指) (图2B)。箭头出现视为刺激呈现, 刺激在屏幕上停留时间为800 ms, 刺激间隔300 ms。实验任务共包括10个组块, 每个组块包含10个序列, 每个序列包含12个不同方向的箭头刺激试次。组块与组块之间被试有充分时间进行休息。在实验开始前, 被试将进行10次练习了解实验程序。对于每个序列中的12个刺激, 共有两种不同的顺序, 对于序列学习组来说, 黑色箭头的呈现顺序是固定的, 将按照1-9-3-1-11-3-11-9-1-3-9-11点钟的方向重复出现。对于随机学习组来说, 每个组块中的120试次的出现顺序为伪随机状态, 每个组块中的顺序都遵循以下限制条件:箭头出现在每个位置的概率相同, 没有直接重复(如1111), 并且排除顺串(如1234)或部分重复(如1212)。参加实验2的40名被试被随机分成两组参加实验, 两组被试的人口学变量不存在差异, 详见表1。
表1 实验2两组被试人口学变量描述统计值和差异检验(M ± SD)
| 变量 | 序列学习组 | 随机学习组 | p |
|---|---|---|---|
| 年龄(岁) | 21.55 ± 1.57 | 21.95 ± 1.88 | 0.469 |
| 身高(cm) | 172.95 ± 9.77 | 171.80 ± 10.35 | 0.720 |
| 体重(kg) | 67.35 ± 13.33 | 66.70 ± 14.81 | 0.885 |
| 受教育年限(年) | 15.90 ± 1.71 | 15.65 ± 1.90 | 0.665 |
| RMT (%) | 43.75 ± 4.72 | 43.80 ± 4.29 | 0.972 |
2.4 数据分析及处理
记录MEP波峰与波谷间的峰峰值作为每次诱发的MEP振幅。两连发(CS-TS)刺激诱发的MEP振幅以该MEP振幅与单脉冲TS振幅的百分比表示(图3)。数值报告以平均值 ± 标准误呈现。
图3
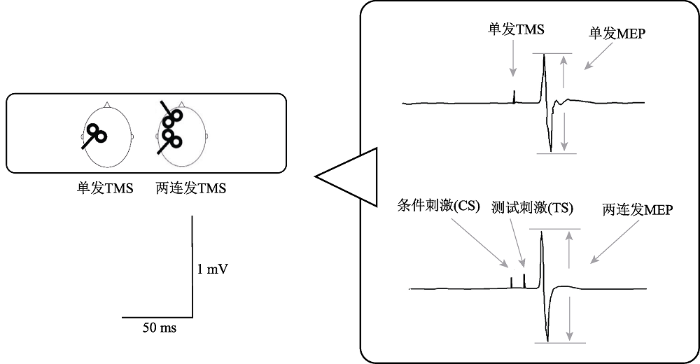
实验1中对DLPFC到M1不同的投射时间间隔下的MEP振幅进行单因素重复测量方差分析考察抑制与兴奋的峰值; 实验2中对行为任务结果进行2(组别) × 2(组块)两因素重复测量方差分析考察两组被试学习效果的差异; 对生理学数据进行2(组别) × 2(前后测) × 5(刺激强度)的三因素重复测量方差分析考察不同条件下单发TMS以及两连发TMS输入输出曲线的差异。数据的检验统计通过SPSS 17.0完成, 显著性判别标准设为p < 0.05。
3 研究结果
3.1 实验1:背外侧前额叶到初级运动皮层投射的时间过程
单因素重复测量方差分析显示不同的时间间隔下双发MEP振幅与单发TS振幅存在显著差异, F (10, 200) = 8.88, p < 0.001, ƞp2 = 0.31 (图4)。采用Bonferrioni法对每个时间间隔下的MEP振幅与单脉冲1 mV的振幅进行多重比较结果显示, 与单脉冲MEP相比, 在时间间隔为10 ms时, MEP振幅显著降低(p = 0.019), 在时间间隔为20 ms时, MEP振幅显著降低(p = 0.021), 而在时间间隔为25 ms时, MEP振幅显著提升(p = 0.047), 其余时间间隔点的统计结果不显著。由于实验需要筛选最大抑制时间点和最大易化时间点, 因此呈现出峰值抑制和峰值易化的10 ms和25 ms将选取为实验2的主要时间点。
图4
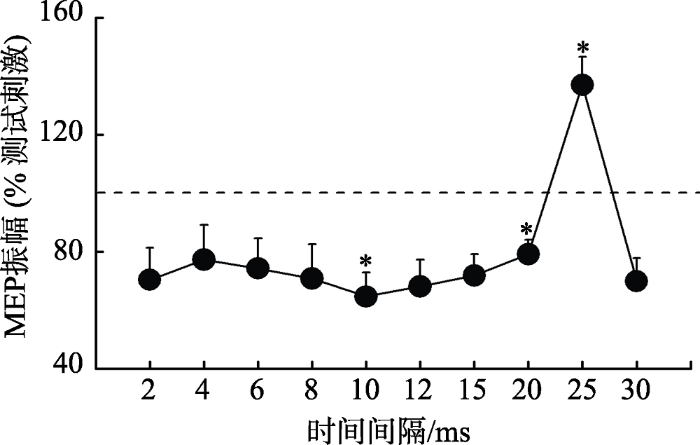
图4
实验1背外侧前额叶到初级运动皮层投射结果
注:时间间隔为第一发刺激(DLPFC)与第二发刺激(M1)之间的时间差, 纵坐标为实际MEP的振幅与单发测试刺激比值后的标准化值。*p < 0.05
3.2 实验2:行为任务反应时间的变化
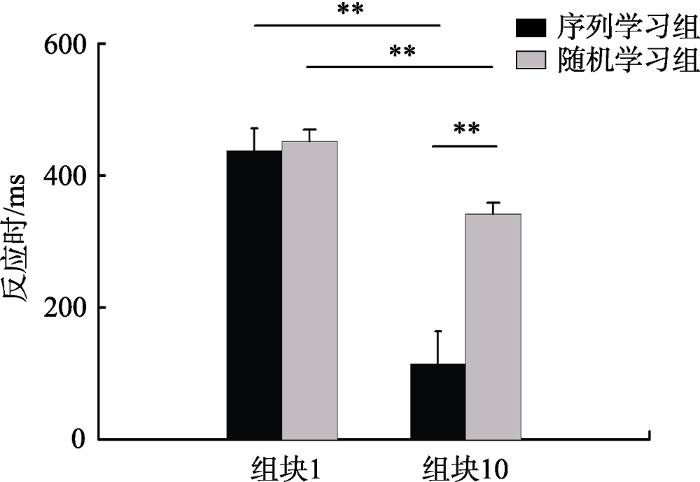
图5显示了两种不同运动学习组别中被试的第一个组块和最后一个组块的平均反应时间。对反应时进行2×2两因素重复测量方差分析显示组别主效应显著, F (1, 38) = 8.33, p = 0.006, ƞp2 = 0.18; 组块主效应显著, F (1, 38) = 138.62, p < 0.001, ƞp2 = 0.79; 组别×组块的交互效应显著, F (1, 38) = 33.49, p < 0.001, ƞp2 = 0.47。简单效应分析显示序列学习组(p < 0.001)与随机学习组(p = 0.018)的反应时下降均显著, 表明由于练习效应的存在, 两组被试组块10的反应时均显著低于组块1的反应时; 在组块10反应时中, 序列学习组与随机学习组存在显著差异(p < 0.001)。
图5
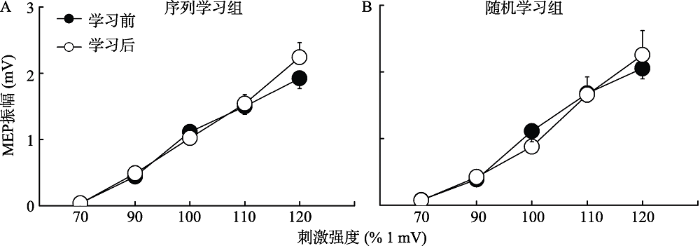
3.3 实验2:不同类型运动学习后单发MEP的IO曲线变化
图6显示了两种不同类型的程序性运动学习前后第一背侧骨间肌的MEP变化。对MEP进行2×2×5三因素重复测量方差分析显示刺激强度主效应显著, F (4, 152) = 140.34, p < 0.001, ƞp2 = 0.79; 前后测主效应不显著, F (1, 38) = 0.17, p = 0.681; 三因素的交互作用不显著, F (4, 152) = 0.11, p = 0.105。
图6
3.4 实验2:不同类型运动学习后两连发MEP的IO曲线变化
根据实验1结果, 选出两个达到兴奋或抑制峰值效应所需时间间隔(10和25 ms)。
在时间间隔为10 ms时, 对MEP进行2×2×5三因素重复测量方差分析显示前后测主效应不显著, F (1, 38) = 0.53, p = 0.472; 刺激强度主效应不显著, F (4, 152) = 1.00, p = 0.411; 三因素交互作用显著, F (4, 152) = 2.61, p = 0.038, ƞp2 = 0.06。随后对前后测和刺激强度进行两因素重复测量方差分析显示, 在序列学习中, 刺激强度主效应不显著, F (4, 76) = 0.98, p = 0.426; 前后测主效应不显著, F (1, 19) = 1.73, p = 0.204; 但前后测×刺激强度交互作用显著, F (4, 76) = 4.24, p = 0.011, ƞp2 = 0.18 (图7A)。事后检验显示在序列学习后, 110%RMT强度下诱发的MEP振幅显著高于学习前(基线水平) (p = 0.002)。在随机学习组中, 两因素方差分析显示刺激强度(F (4, 76) = 0.57, p = 0.683)或前后测组别(F (1, 19) = 0.12, p = 0.736)或前后测×刺激强度的交互作用(F (4, 76) = 0.77, p = 0.551)均不显著(图7B)。
图7
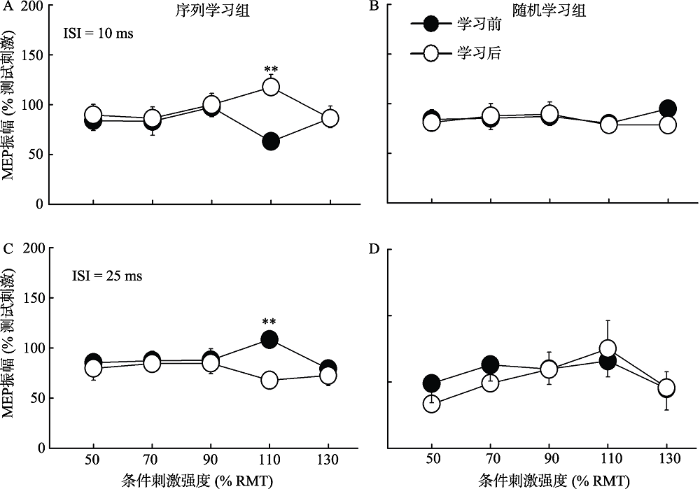
在时间间隔为25 ms时, 对MEP进行2×2×5三因素重复测量方差分析, 结果发现前后测主效应不显著, F (1, 38) = 1.08, p = 0.304; 刺激强度主效应显著, F (4, 152) = 3.86, p = 0.005, ƞp2 = 0.09; 三因素交互作用显著, F (4, 152) = 2.48, p = 0.046, ƞp2 = 0.06。随后进行的两因素重复测量方差分析显示, 在序列学习组中, 刺激强度(F (4, 76) = 1.07, p = 0.376)和前后测(F (1, 19) = 3.00, p = 0.099)主效应均不显著, 但前后测×刺激强度交互作用显著, F (4, 76) = 3.29, p = 0.015, ƞp2 = 0.15 (图7C)。事后检验显示在序列学习后, 110% RMT强度下诱发的MEP振幅显著低于学习前(基线水平) (p < 0.001)。在随机学习组中, 两因素方差分析显示刺激强度主效应显著, F (4, 76) = 3.23, p = 0.017, ƞp2 = 0.15。事后比较显示50% RMT与90% RMT (p = 0.041), 50% RMT与110% RMT (p = 0.011), 110% RMT与130% RMT (p = 0.033)之间存在显著差异; 但是前后测(F (1, 19) = 0.09, p = 0.772)或前后测×刺激强度的交互作用(F (4, 76) = 0.65, p = 0.626)均不显著(图7D)。
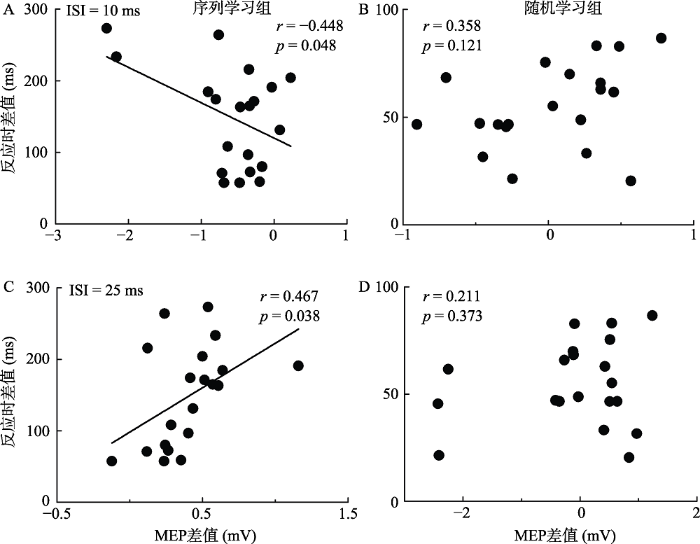
3.5 行为数据与生理数据的相关
图8显示了两种不同运动学习组别中被试的第一个组块和最后一个组块的平均反应时间之差与生理学指标MEP之差的相关统计结果。MEP之差为正值说明学习后运动皮层兴奋性下降, 反之则上升。通过相关分析发现, 被试在时间间隔为10 ms时, 序列学习组的运动皮层兴奋性上升幅度越大, 所用的反应时下降也越大, r= -0.448, p = 0.048(图8A); 在时间间隔为25 ms时, 序列学习组的运动皮层兴奋性下降幅度越大, 所用的反应时下降幅度也越大, r = 0.467, p = 0.038 (图8C)。随机学习组在两个时间间隔中均未表现出显著相关性(10 ms时, p = 0.121, 图8B; 25 ms时, p = 0.373, 图8D)。
图8
4 讨论
本研究采用SRTT和TMS, 探究左侧DLPFC到M1的联通性在两种不同的程序性运动学习中是否有差异。通过行为和电生理数据, 我们发现(1)序列学习组的学习效果更好, 随着学习的进行, 反应时显著下降。(2)左侧DLPFC到M1的投射在两个时间节点上分别表现出显著的抑制(10 ms)和易化连接极值(25 ms)。(3)单发TMS产生的MEP在序列学习和随机学习两种学习条件下均未发生改变; 而两连发的MEP显示, 在110% RMT的输入强度下, DLPFC-M1联通性在序列条件下发生改变, 且与行为数据相关, 而在随机条件下未发生改变。
4.1 DLPFC到M1投射的抑制性及易化性神经通路
结果发现, 从DLPFC到M1有早期的抑制峰值和晚期的易化峰值(图4)。这表明第一发的CS通过激活前额叶皮层而抑制或增强了初级运动皮层对第二发TS的反应。存在的这两个投射时间点可能是因为从特定脑功能区到M1的大脑纤维通路长度不同和相关神经元的阈值不同等原因造成的(Wedeen et al., 2012)。早期出现的10 ms抑制峰值可能是突触信号从DLPFC直接传递到M1的最佳时间, 这取决于来自前额叶或来自于 M1的潜在白质(即上纵束)对皮质直接投射的激活(Koch et al., 2007)。另外, 抑制了DLPFC的兴奋性后, M1的皮层兴奋性提升; 而易化了DLPFC的兴奋性后, M1的兴奋性下降(Cao et al., 2018)。这种双向的反作用关系表明了DLPFC与M1存在着抑制的投射通路, 与本研究的发现一致。对于后期出现的25 ms的易化峰值, 可能的解释是输入信号的传递并不是完全从DLPFC直接传到M1, 而是途经背侧前运动区等其他脑区, 它们扮演着“中转站”角色, 形成多突触回路传递。大多数来自于DLPFC的皮质投射会经由前运动皮质, 到达初级运动皮质, 因此在25 ms的时间节点存在易化效应(Koch & Rothwell, 2009)。
除了在10 ms和25 ms时间间隔下分别达到抑制和易化的顶点, 在2、4、6、8、15、20 ms及30 ms的时间间隔中MEP同样呈现出抑制的投射状态。与M1相连的突触投射中, 易化效果只出现在较少的时间间隔下并且对刺激强度的敏感度要求较高, 而在大多数的时间间隔和刺激强度中则呈现出抑制的投射, 这与Koch等人先前研究结果一致(Koch et al., 2007)。抑制投射的主导性原因至今尚不明确。一个可能的解释是易化的连接是区域范围内少数通路之间的投射连接, 而抑制投射则是大面积、更广泛普遍的存在。TMS作为焦点刺激法, 每个线圈的磁刺激只能施加到目标区域中具体的某一焦点, 因此更容易探知到更广泛存在的抑制连接(Hanajima et al., 2001)。
4.2 DLPFC在不同程序性运动学习中的差别性作用
研究发现与随机学习组相比, 序列学习组DLPFC-M1通路的兴奋性有不同的变化, 特别是在CS强度为110% RMT时表现最强(图7A, C)。基于以往的研究, 可能的解释是相比于阈下刺激, 阈上刺激能够更好地诱发额叶和M1之间的功能性作用与连接(Jacinta et al., 2010; Koch et al., 2007), 而当阈上刺激的强度过高(130% RMT)时, 反而会抑制该通路中的突触传递, 从而抑制测试刺激诱发的MEP (Uehara, Morishita, Kubota, & Funase, 2013)。
实验2结果发现序列学习组可以在两个最佳时间点改变DLPFC-M1的兴奋性, 说明序列学习和随机学习在DLPFC-M1投射上存在差异。在序列学习任务中, 屏幕上出现的信号很快到达前额叶区域并获得加工, 经过足够试次的训练, DLPFC进一步激活, 并向运动皮质发送促通信号, 导致DLPFC-M1的联通性在10 ms的间隔时间下比学习前显著增强(图7A)。而在25 ms的时间间隔点下, 两连发MEP在序列学习后显著下降(图7C), 同10 ms时间点的MEP一样显示出了与基线状态相反的变化趋势。这种同步的反转展示出了学习产生的效应对于脑区联通性的影响, 学习后记忆的痕迹带来的独特、持久的表征作用于大脑网络, 产生可塑性的变化进而产生反应时的变化(张剑心, 汤旦, 查德华, 黄建平, 刘电芝, 2016)。前人采用fMRI方法也报告了序列学习后DLPFC和M1的静息态功能连接(resting-state functional connectivity)增强的结果(Steel et al., 2016)。
序列学习被认为是通过经验的积累来完成学习(Mayor-Dubois et al., 2016)。其与随机学习的主要区别在于受试者需要通过激活DLPFC调用空间注意力伴随意识模式发现并掌握规律, 来完成记忆序列和决策的过程(杨叶红, 王树明, 2018)。神经影像学研究证实左侧DLPFC在运动学习中被激活(Toni et al., 1998) 。对于序列学习任务, 前人研究结果表明在按键学习任务中对DLPFC的脉冲刺激显著增强了被试M1的兴奋性, 这与我们在序列学习组10 ms两连发TMS的结果是一致的(Hasan et al., 2013; Lam et al., 2015)。Willingham等人(2002)认为DLPFC可能传递一种与意识的注意方向平行的功能即有意识的选择性反应。这种在目标出现之前的有意识的选择会降低序列学习条件下的反应时。
值得一提的是, 尽管受试者没有提前知道序列的存在, 但在本研究中, 序列学习组的受试者均在运动学习任务结束后成功地回忆起了包含12个数字的整个序列顺序。也就是说, 左侧DLPFC区域的激活与通过观察线索的出现进行序列搜索的过程有关。DLPFC参与了序列搜索的过程, 这些意识参与并且再认的过程是目标导向模式搜索中的内隐过程, 进而激活背外侧前额叶脑区(Fatma, Thorsten, Christof, & John-Dylan, 2012)。我们同样对生理数据和行为数据进行了相关性的分析, 结果显示在序列学习组中, 无论是10 ms还是25 ms, 被试在第一个组块与最后一个组块的反应时差值和学习前后两连发TMS呈现显著的相关性(图8A, C), 表现出DLPFC-M1的联通性与行为任务中的学习效果提升关系密切, 进一步表明左侧DLPFC的激活水平与受试者序列学习的能力有关。
4.3 运动认知网络在运动学习中的重要性
我们发现两种不同的运动学习任务干预约20分钟后, 单发运动诱发电位的IO曲线没有任何变化(图6), 相反, 两连发MEP的IO曲线在序列顺序组中发生了显著性变化(图7A, C), 表明序列学习对DLPFC-M1通路的兴奋性有一定的影响。Jueptner等人在1997年的一项脑成像研究中也报告了类似的结果, 这些受试者被要求用8个手指对一个类似的序列学习任务重复训练, 并对训练前后受试者的运动兴奋性进行比较。结果表明在训练的早期阶段, 短期的、同组块内的可塑性并没有显著性变化(Jueptner et al., 1997), 这个结果也得到了其他研究的支持(Grafton, Hazeltine, & Ivry, 1995; Stefan et al., 2006)。这些研究表明短期的运动学习激活了相关的大脑网络, 但是没有特别地对M1有显著性的激活和兴奋性的提升。
已有的很多文献都对M1的抑制和促进效应进行过探究, 其中条件刺激来自于同侧小脑(Ugawa et al., 1991)或施加于前额叶区等脑功能区(Cao et al., 2018; Civardi, Cantello, Asselman, & Rothwell, 2001)。这些通路和不同脑区之间的联通性表明大脑皮层的信息传递和组织具有全局性, 意识的产生和动作的执行必须在全脑建立长时程的工作平台, 而不仅仅局限于某一单一的脑功能区。具有不同生理特性的特定神经元群分布在大脑皮层网络的各个节点上, 并与远近各处的其他神经元形成极其复杂的连接(Oh et al., 2014)。前人的这些研究主要探讨了静息状态下DLPFC和M1之间的联系与投射, 而没有分析运动学习对两个功能脑区联通性的影响。本研究加入了程序性运动学习任务, 进一步发现了DLPFC-M1的联通性对于提升运动学习中的表现有显著的作用, 从而说明DLPFC-M1通路在序列运动学习中的重要性与因果性。通过了解DLPFC到M1的投射在序列和随机学习中的差别性作用, 能让我们更多地了解 DLPFC脑区在其众多功能中的分化与合作。探究人类大脑各功能区之间的协同合作和网络工作模式依旧是揭示人类日常心理活动的重要课题。
5 结论
本研究通过连续反应时任务和经颅磁刺激法的结合对左侧背外侧前额叶在程序性运动学习中的作用进行探究, 行为学结果发现序列学习组的学习效果更佳; 电生理学结果发现, 两组学习前、后M1的运动诱发电位均未发生改变; 在最佳时间投射点、适当刺激强度下, 序列学习组DLPFC-M1联通性发生改变, 且与学习成绩相关, 而随机学习组没有改变。结果表明, DLPFC和M1间联通性的增强可能是序列学习成绩更佳的重要原因, 这一研究从电生理角度为DLPFC在运动学习中的作用提供了重要证据。
参考文献
Inhibitory and facilitatory connections from dorsolateral prefrontal to primary motor cortex in healthy humans at rest-an rTMS study
DOI:10.1016/j.neulet.2018.09.032
URL
PMID:30243883
[本文引用: 3]

The human motor system consists of several divisions in the frontal lobes. The physiological function of projections from the dorsolateral prefrontal cortex (DLPFC) to the primary motor cortex (M1) remains elusive. Here, we introduce theta burst stimulation (TBS)-based protocols to target inhibitory and facilitatory connections in the DLPFC-M1 network.
Transcranial magnetic stimulation can be used to test connections to primary motor areas from frontal and medial cortex in humans
DOI:10.1006/nimg.2001.0918
URL
PMID:11707100
[本文引用: 1]
Surface EMG responses (MEPs) were recorded from the relaxed first dorsal interosseous (FDI) of 16 normal subjects following transcranial magnetic stimulation (TMS) over the hand area of the primary motor cortex. These test responses were conditioned by a subthreshold stimulus applied 2-15 ms beforehand over a range of anterior or medial sites. Stimuli applied 3-5 cm anterior to the hand motor area (site A) or 6 cm anterior to the vertex on the nasion-inion line (site B) inhibited the test responses at short latency. The largest effect was seen when the interstimulus interval was 6 ms and the intensity of the conditioning stimulus was equal to 0.9x active motor threshold (AMT) at the hand area. Increasing the intensity to 1.2x AMT produced facilitation. Suppression of surface EMG responses was mirrored in the behavior of single motor units. Conditioning stimuli had no effect on responses evoked in the active FDI muscle by transcranial electric stimulation of motor cortex nor on forearm flexor H reflexes even though MEPs in the same muscle were suppressed at appropriate interstimulus intervals. We conclude that low-intensity TMS over presumed premotor areas of frontal cortex can engage corticocortical connections to the primary motor hand area.
Sequence learning
DOI:10.1016/s1364-6613(98)01202-9
URL
PMID:21227209
[本文引用: 1]
The ability to sequence information is fundamental to human performance. When subjects are asked to respond to one of several possible spatial locations of a stimulus, reaction times and error rates decrease when the target follows a sequence. In this article, we review the numerous theoretical and methodological perspectives that have been used to study sequence learning. The opportunity now exists to integrate evidence from different domains of cognitive science to begin to provide a comprehensive account of sequence learning. We suggest that subjects can learn sequences based on different information in a hierarchical representation, including either sequences of stimuli or sequences of responses. This learning can occur both with and without explicit awareness of the sequence. Multiple modes of learning exist and are subserved by different neural circuits.
Changes in functional connectivity support conscious object recognition
DOI:10.1016/j.neuroimage.2012.07.056
URL
PMID:22877578
[本文引用: 1]
What are the brain mechanisms that mediate conscious object recognition? To investigate this question, it is essential to distinguish between brain processes that cause conscious recognition of a stimulus from other correlates of its sensory processing. Previous fMRI studies have identified large-scale brain activity ranging from striate to high-level sensory and prefrontal regions associated with conscious visual perception or recognition. However, the possible role of changes in connectivity during conscious perception between these regions has only rarely been studied. Here, we used fMRI and connectivity analyses, together with 120 custom-generated, two-tone, Mooney images to directly assess whether conscious recognition of an object is accompanied by a dynamical change in the functional coupling between extrastriate cortex and prefrontal areas. We compared recognizing an object versus not recognizing it in 19 naïve subjects using two different response modalities. We find that connectivity between the extrastriate cortex and the dorsolateral prefrontal cortex (DLPFC) increases when objects are consciously recognized. This interaction was independent of the response modality used to report conscious recognition. Furthermore, computing the difference in Granger causality between recognized and not recognized conditions reveals stronger feedforward connectivity than feedback connectivity when subjects recognized the objects. We suggest that frontal and visual brain regions are part of a functional network that supports conscious object recognition by changes in functional connectivity.
Functional and effective connectivity: A review
DOI:10.1089/brain.2011.0008
URL
PMID:22432952
[本文引用: 1]
Over the past 20 years, neuroimaging has become a predominant technique in systems neuroscience. One might envisage that over the next 20 years the neuroimaging of distributed processing and connectivity will play a major role in disclosing the brain's functional architecture and operational principles. The inception of this journal has been foreshadowed by an ever-increasing number of publications on functional connectivity, causal modeling, connectomics, and multivariate analyses of distributed patterns of brain responses. I accepted the invitation to write this review with great pleasure and hope to celebrate and critique the achievements to date, while addressing the challenges ahead.
Neuron activity related to short-term memory
DOI:10.1126/science.173.3997.652
URL
PMID:4998337
[本文引用: 1]
Nerve cells in the monkey's prefrontal cortex and nucleus medialis dorsalis of the thalamus show changes of firing frequency associated with the performance of a delayed response test. Most cells increase firing during the cue presentation period or at the beginning of the ensuing delay; spike discharge highler than that in intertrial periods is present in some cells throughout the delay. These changes are interpreted as suggestive evidence of a role of frontothalamic circuits in the attentive process involved in short-term memory
Preserved learning of novel information in amnesia: Evidence for multiple memory systems
DOI:10.1016/0278-2626(88)90002-4
URL
PMID:3401382
[本文引用: 1]
Four of five patients with marked global amnesia, and others with new learning impairments, showed normal processing facilitation for novel stimuli (nonwords) and/or for familiar stimuli (words) on a word/nonword (lexical) decision task. The data are interpreted as a reflection of the learning capabilities of in-line neural processing stages with multiple, distinct, informational codes. These in-line learning processes are separate from the recognition/recall memory impaired by amygdalohippocampal/dosomedial thalamic damage, but probably supplement such memory in some tasks in normal individuals. Preserved learning of novel information seems incompatible with explanations of spared learning in amnesia that are based on the episodic/semantic or memory/habit distinctions, but is consistent with the procedural/declarative hypothesis.
Implicit learning in patients with Alzheimer's disease
DOI:10.1055/s-2007-1014490
URL
PMID:2339182
[本文引用: 1]
We examined implicit memory using priming and procedural learning tasks in patients with probable Dementia-Alzheimer's Type (DAT) to examine whether priming and procedural processes could be dissociated and whether task specificity was a factor in DAT patient performance. Priming was tested using a word recognition paradigm (perceptual priming) and by repeated administrations of a fragmented objects test (long term priming). Procedural learning was tested using repeated and random sequences on a choice serial reaction time task and by repeated administration of a puzzle map of the United States. DAT patients were compared to hospitalized depressed patients, patients suffering from Progressive Supranuclear Palsy (PSP), and normal controls. We found that DAT patients demonstrated marginal but significant implicit learning on both procedural learning and perceptual priming tasks. DAT patients performed relatively better on the procedural learning task than a perceptual priming task compared to PSP patients, suggesting that priming of meaningful stimuli is subserved by cortical structures whereas procedural motor responses to simple serial visual stimulus patterns can be maintained by subcortical systems. Furthermore, our findings suggest that priming and procedural processes can be dissociated and that task specificity is a factor in interpreting the results of implicit learning paradigms in DAT patients. The implications of these results for models of knowledge representation and memory processes as well as the way they can serve as models for testing nootropic drug effects are discussed.
Functional imaging of procedural motor learning: Relating cerebral blood flow with individual subject performance
DOI:10.1002/hbm.460010307
URL
PMID:24578042
[本文引用: 1]
Changes of local synaptic activity during acquisition of a visuomotor skill were examined with positron emission tomography (PET) imaging of regional cerebral blood flow (rCBF). Eight subject learned the pursuit rotor task, a predictable tracking task, during three sequential PET scans (day 1). Subjects returned 2 days later and repeated the three pursuit trials and PET scans (day 2) after completing an extensive practice session. Control scans without movement bracketed the pursuit trials on both days to rule out time effects unrelated to motor skill learning. PET images were transformed to a common stereotaxic space using matched magnetic resonance imaging (MRI) scans. Group learning effects were determined by a repeated measures multivariate analysis of variance (ANOVA). During motor skill acquisition (day 1), increases of synaptic activity were identified in cortical motor areas and cerebellum, supporting the hypothesis that procedural motor learning occurs in motor execution areas. During long-term practice (day 2), changes were limited to the bilateral putamen, bilateral parietal cortex, and left premotor cortex. To characterize differences in the rate of learning between subjects, each subject's performance data from day 1 was fit with a power function. The exponents were correlated with rCBF data on a pixel-by-pixel basis. Rapid skill acquisition was associated with increasing rCBF in premotor, prefrontal, and cingulate areas, and decreasing rCBF in visual processing areas located in the temporal and occipital cortex. This pattern in fast learners may reflect a more rapid shift from a visually guided strategy (accessing perceptual areas) to an internally generated model (accessing premotor and prefrontal areas). © 1994 Wiley-Liss, Inc.
Transcranial magnetic stimulation and the human brain
DOI:10.1038/35018000
URL
PMID:10910346
[本文引用: 2]
Transcranial magnetic stimulation (TMS) is rapidly developing as a powerful, non-invasive tool for studying the human brain. A pulsed magnetic field creates current flow in the brain and can temporarily excite or inhibit specific areas. TMS of motor cortex can produce a muscle twitch or block movement; TMS of occipital cortex can produce visual phosphenes or scotomas. TMS can also alter the functioning of the brain beyond the time of stimulation, offering potential for therapy.
Interhemispheric facilitation of the hand motor area in humans
DOI:10.1111/j.1469-7793.2001.0849h.x
URL
PMID:11251064
[本文引用: 1]
1. We investigated interhemispheric interactions between the human hand motor areas using transcranial cortical magnetic and electrical stimulation. 2. A magnetic test stimulus was applied over the motor cortex contralateral to the recorded muscle (test motor cortex), and an electrical or magnetic conditioning stimulus was applied over the ipsilateral hemisphere (conditioning motor cortex). We investigated the effects of the conditioning stimulus on responses to the test stimulus. 3. Two effects were elicited at different interstimulus intervals (ISIs): early facilitation (ISI = 4-5 ms) and late inhibition (ISI &gt; or = 11 ms). 4. The early facilitation was evoked by a magnetic or anodal electrical conditioning stimulus over the motor point in the conditioning hemisphere, which suggests that the conditioning stimulus for early facilitation directly activates corticospinal neurones. 5. The ISIs for early facilitation taken together with the time required for activation of corticospinal neurones by I3-waves in the test hemisphere are compatible with the interhemispheric conduction time through the corpus callosum. Early facilitation was observed in responses to I3-waves, but not in responses to D-waves nor to I1-waves. Based on these results, we conclude that early facilitation is mediated through the corpus callosum. 6. If the magnetic conditioning stimulus induced posteriorly directed currents, or if an anodal electrical conditioning stimulus was applied over a point 2 cm anterior to the motor point, then we observed late inhibition with no early facilitation. 7. Late inhibition was evoked in responses to both I1- and I3-waves, but was not evoked in responses to D-waves. The stronger the conditioning stimulus was, the greater was the amount of inhibition. These results are compatible with surround inhibition at the motor cortex.
Muscle and timing-specific functional connectivity between the dorsolateral prefrontal cortex and the primary motor cortex
DOI:10.1162/jocn_a_00338
URL
PMID:23249357
[本文引用: 2]
The pFC has a crucial role in cognitive control, executive function, and sensory processing. Functional imaging, neurophysiological, and animal studies provide evidence for a functional connectivity between the dorsolateral pFC (DLPFC) and the primary motor cortex (M1) during free choice but not instructed choice selection tasks. In this study, twin coil, neuronavigated TMS was used to examine the precise timing of the functional interaction between human left DLPFC and ipsilateral M1 during the execution of a free/specified choice selection task involving the digits of the right hand. In a thumb muscle that was not involved in the task, a conditioning pulse to the left DLPFC enhanced the excitability of the ipsilateral M1 during free selection more than specified selection 100 msec after presentation of the cue; the opposite effect was seen at 75 msec. However, the difference between free and externally specified conditions disappeared when a task-specific muscle was investigated. In this case, the influence from DLPFC was dominated by task involvement rather than mode of selection, suggesting that other processes related to movement execution were also operating. Finally, we show that the effects were spatially specific because they were absent when an adjacent area of DLPFC was stimulated. These results reveal temporally and spatially selective interactions between BA 46 and M1 that are both task and muscle specific.
Central mechanisms of motor skill learning
DOI:10.1016/s0959-4388(02)00307-0
URL
PMID:12015240
[本文引用: 2]
Recent studies have shown that frontoparietal cortices and interconnecting regions in the basal ganglia and the cerebellum are related to motor skill learning. We propose that motor skill learning occurs independently and in different coordinates in two sets of loop circuits: cortex-basal ganglia and cortex-cerebellum. This architecture accounts for the seemingly diverse features of motor learning.
Functional specificity of human premotor-motor cortical interactions during action selection
DOI:10.1111/j.1460-9568.2007.05795.x
URL
PMID:17868374
[本文引用: 2]
Functional connections between dorsal premotor cortex (PMd) and primary motor cortex (M1) have been revealed by paired-pulse transcranial magnetic stimulation (TMS). We tested if such connections would be modulated during a cognitive process (response selection) known to rely on those circuits. PMd-M1 TMS applied 75 ms after a cue to select a manual response facilitated motor-evoked potentials (MEPs). MEPs were facilitated at 50 ms in a control task of response execution, suggesting that PMd-M1 interactions at 75 ms are functionally specific to the process of response selection. At 100 ms, PMd-M1 TMS delayed choice reaction time (RT). Importantly, the MEP (at 75 ms) and the RT (at 100 ms) effects were correlated in a way that was hand-specific. When the response was made with the M1-contralateral hand, MEPs correlated with slower RTs. When the response was made with the M1-ipsilateral hand, MEPs correlated with faster RTs. Paired-pulse TMS confined to M1 did not produce these effects, confirming the causal influence of PMd inputs. This study shows that a response selection signal evolves in PMd early during the reaction period (75-100 ms), impacts on M1 and affects behaviour. Such interactions are temporally, anatomically and functionally specific, and have a causal role in choosing which movement to make.
Motor sequence learning: A study with positron emission tomography
We have used positron emission tomography to study the functional anatomy of motor sequence learning. Subjects learned sequences of keypresses by trial and error using auditory feedback. They were scanned with eyes closed under three conditions: at rest, while performing a sequence that was practiced before scanning until overlearned, and while learning new sequences at the same rate of performance. Compared with rest, both sequence tasks activated the contralateral sensorimotor cortex to the same extent. Comparing new learning with performance of the prelearned sequence, differences in activation were identified in other areas. (1) Prefrontal cortex was only activated during new sequence learning. (2) Lateral premotor cortex was significantly more activated during new learning, whereas the supplementary motor area was more activated during performance of the prelearned sequence. (3) Activation of parietal association cortex was present during both motor tasks, but was significantly greater during new learning. (4) The putamen was equally activated by both conditions. (5) The cerebellum was activated by both conditions, but the activation was more extensive and greater in degree during new learning. There was an extensive decrease in the activity of prestriate cortex, inferotemporal cortex, and the hippocampus in both active conditions, when compared with rest. These decreases were significantly greater during new learning. We draw three main conclusions. (1) The cerebellum is involved in the process by which motor tasks become automatic, whereas the putamen is equally activated by sequence learning and retrieval, and may play a similar role in both. (2) When subjects learn new sequences of motor actions, prefrontal cortex is activated. This may reflect the need to generate new responses. (3) Reduced activity of areas concerned with visual processing, particularly during new learning, suggests that selective attention may involve depressing the activity of cells in modalities that are not engaged by the task.
Anatomy of motor learning. I. Frontal cortex and attention to action
DOI:10.1152/jn.1997.77.3.1313
URL
PMID:9084599
[本文引用: 1]
We used positron emission tomography to study new learning and automatic performance in normal volunteers. Subjects learned sequences of eight finger movements by trial and error. In a previous experiment we showed that the prefrontal cortex was activated during new learning but not during during automatic performance. The aim of the present experiment was to see what areas could be reactivated if the subjects performed the prelearned sequence but were required to pay attention to what they were doing. Scans were carried out under four conditions. In the first the subjects performed a prelearned sequence of eight key presses; this sequence was learned before scanning and was practiced until it had become overlearned, so that the subjects were able to perform it automatically. In the second condition the subjects learned a new sequence during scanning. In a third condition the subjects performed the prelearned sequence, but they were required to attend to what they were doing; they were instructed to think about the next movement. The fourth condition was a baseline condition. As in the earlier study, the dorsal prefrontal cortex and anterior cingulate area 32 were activated during new learning, but not during automatic performance. The left dorsal prefrontal cortex and the right anterior cingulate cortex were reactivated when subjects paid attention to the performance of the prelearned sequence compared with automatic performance of the same task. It is suggested that the critical feature was that the subjects were required to attend to the preparation of their responses. However, the dorsal prefrontal cortex and the anterior cingulate cortex were activated more when the subjects learned a new sequence than they were when subjects simply paid attention to a prelearned sequence. New learning differs from the attention condition in that the subjects generated moves, monitored the outcomes, and remembered the responses that had been successful. All these are nonroutine operations to which the subjects must attend. Further analysis is needed to specify which are the nonroutine operations that require the involvement of the dorsal prefrontal and anterior cingulate cortex.
Inside the brain of an elite athlete: the neural processes that support high achievement in sports
DOI:10.1038/nrn2672
URL
PMID:19571792
[本文引用: 1]
Events like the World Championships in athletics and the Olympic Games raise the public profile of competitive sports. They may also leave us wondering what sets the competitors in these events apart from those of us who simply watch. Here we attempt to link neural and cognitive processes that have been found to be important for elite performance with computational and physiological theories inspired by much simpler laboratory tasks. In this way we hope to inspire neuroscientists to consider how their basic research might help to explain sporting skill at the highest levels of performance.
Focal stimulation of the posterior parietal cortex increases the excitability of the ipsilateral motor cortex
DOI:10.1523/JNEUROSCI.0598-07.2007
URL
PMID:17581969
[本文引用: 4]
Paired-pulse transcranial magnetic stimulation (TMS) has been applied as a probe to test functional connectivity within distinct cortical areas of the human motor system. Here, we tested the interaction between the posterior parietal cortex (PPC) and ipsilateral motor cortex (M1). A conditioning TMS pulse over the right PPC potentiates motor evoked-potentials evoked by a test TMS pulse over the ipsilateral motor cortex, with a time course characterized by two phases: an early peak at 4 ms interstimulus interval (ISI) and a late peak at 15 ms ISI. Activation of this facilitatory pathway depends on the intensity of stimulation, because the effects are induced with a conditioning stimulus of 90% resting motor threshold but not at lower or higher intensities. Similar results were obtained testing the ipsilateral interaction in the left hemisphere with a slightly different time course. In control experiments, we found that activation of this facilitatory pathway depends on the direction of induced current in the brain and is specific for stimulation of the caudal part of the inferior parietal sulcus (cIPS) site, because it is not observed for stimulation of adjacent scalp sites. Finally, we found that by using poststimulus time histogram analysis of single motor unit firing, the PPC conditioning increases the excitability of ipsilateral M1, enhancing the relative amount of late I wave input recruited by the test stimulus over M1, suggesting that such interaction is mediated by specific interneurons in the motor cortex. The described facilitatory connections between cIPS and M1 may be important in a variety of motor tasks and neuropsychiatric disorders.
TMS investigations into the task-dependent functional interplay between human posterior parietal and motor cortex
DOI:10.1016/j.bbr.2009.03.023
URL
PMID:19463695
[本文引用: 1]
Transcranial magnetic stimulation (TMS) can be used in two different ways to investigate the contribution of cortical areas involved in grasp/reach movements in humans. It can produce &quot;virtual lesions&quot; that interfere with activity in particular cortical areas at specific times during a task, or it can be used in a twin coil design to test the excitability of cortical projections to M1 at different times during a task. The former method has described how cortical structures such as the ventral premotor cortex (PMv), dorsal premotor cortex (PMd) and the anterior intraparietal sulcus (aIPS) are important for specific aspects of reaching, grasping and lifting objects. In the latter method, a conditioning stimulus (CS) is first used to activate putative pathways to the motor cortex from, for example, posterior parietal cortex (PPC) or PMd, while a second, test stimulus (TS), delivered over the primary motor cortex a few ms later probes any changes in excitability that are produced by the input. Thus changes in the effectiveness of the conditioning pulse give an indication of how the excitability of the connection changes over time and during a specific task. Here we review studies describing the time course of operation of parallel intracortical circuits and cortico-cortical connections between the PMd, PMv, PPC and M1, thus demonstrating that functional interplay between these areas and the primary motor cortices is not fixed, but can change in a highly task-, condition- and time-dependent manner.
Procedural motor learning in Parkinson’s disease
DOI:10.1007/s002210100871
URL
PMID:11810137
[本文引用: 1]
We have been investigating motor control and learning in parkinsonian subjects. In the current study, we sought to explore the existence of deficits in procedural motor learning, which is a form of implicit motor learning where skill improves over repetitive blocks of trials. We sought to determine, in particular, whether any such deficit is accentuated during specific types or phases of learning. We would expect that those specific learning tasks would require the greatest participation of the basal ganglia. Numerous studies have found that Parkinson's disease (PD) patients may show deficits in learning. Combined with information about basal ganglia neuronal connections and activity, this led some investigators to suggest that one of the key functions of the basal ganglia is to facilitate learning. To investigate these learning deficits, we used a robotic device to generate conservative force fields that disturbed the subjects' arm movements, thereby generating a &quot;virtual mechanical environment&quot; that subjects learned to manipulate. Movements were successively grouped into blocks comprising five different conditions: motor performance, early learning, late learning, negative transfer, and aftereffect motor performance. Our results with eight right-handed PD subjects and nine age-matched controls showed a relative decrease in the rate of learning for the PD patients in all blocks, but greater differences emerged between groups during novelty phases of learning. In particular, the difference in performance during the negative transfer condition reached statistical significance, suggesting that the basal ganglia might be a key center for &quot;switching&quot; motor patterns. Our results support the hypothesis that deficiencies in procedural motor learning are characteristic of PD. They add to existing evidence which has suggested a key role for the basal ganglia when new sensorimotor mappings are required by novel task environments. Better understanding of these deficits should facilitate the rehabilitation of PD patients.
Assessment of effective connectivity and plasticity with dual-coil transcranial magnetic stimulation
DOI:10.1016/j.brs.2016.02.010
URL
PMID:27207765
[本文引用: 1]
Understanding how different brain regions interact with one another is at the heart of current endeavors in cognitive and basic neuroscience. Unlike most neuroimaging techniques, transcranial magnetic stimulation (TMS) allows the establishment of causal relationships in the study of the functional architecture of the human brain. While this tool is increasingly used to probe the functional and causal nature of the associations between brain regions, a comprehensive guide documenting the various existing stimulation protocols is currently lacking, limiting its use.
Effects of age on motor learning and prefrontal-motorcortical excitability
Direct demonstration of interhemispheric inhibition of the human motor cortex produced by transcranial magnetic stimulation
DOI:10.1007/s002210050648
URL
PMID:10090664
[本文引用: 1]
Electromyographic (EMG) responses evoked in hand muscles by a magnetic test stimulus over the motor cortex can be suppressed if a conditioning stimulus is applied to the opposite hemisphere 6-30 ms earlier. In order to define the mechanism and the site of action of this inhibitory phenomenon, we recorded descending volleys produced by the test stimulus through high cervical, epidural electrodes implanted for pain relief in three conscious subjects. These could be compared with simultaneously recorded EMG responses in hand muscles. When the test stimulus was given on its own it evoked three waves of activity (I-waves) in the spinal cord, and a small EMG response in the hand. A prior conditioning stimulus to the other hemisphere suppressed the size of both the descending spinal cord volleys and the EMG responses evoked by the test stimulus when the interstimulus interval was greater than 6 ms. In the spinal recordings, the effect was most marked for the last I-wave (I3), whereas the second I2-wave was only slightly inhibited, and the first I-wave (I1) was not inhibited at all. We conclude that transcranial stimulation over the lateral part of the motor cortex of one hemisphere can suppress the excitability of the contralateral motor cortex.
The contribution of primary motor cortex is essential for probabilistic implicit sequence learning: Evidence from theta burst magnetic stimulation
DOI:10.1162/jocn.2009.21208
URL
PMID:19301999
[本文引用: 2]
Theta burst transcranial magnetic stimulation (TBS) is considered to produce plastic changes in human motor cortex. Here, we examined the inhibitory and excitatory effects of TBS on implicit sequence learning using a probabilistic serial reaction time paradigm. We investigated the involvement of several cortical regions associated with implicit sequence learning by examining probabilistic sequence learning in five age- and IQ-matched groups of healthy participants following continuous inhibitory TBS over primary motor cortex (M1); or the supplementary motor area (SMA) or dorsolateral prefrontal cortex (DLPFC) or following intermittent excitatory TBS of M1; or after sham TBS. Relative to sham TBS, probabilistic sequence learning was abolished by inhibitory TBS over M1, demonstrating that this region is critical for implicit motor sequence learning. Sequence learning was not significantly affected by inhibitory TBS over the SMA, DLPFC or excitatory TBS over M1. These results demonstrate that the M1 mediates implicit sequence learning.
The use of transcranial magnetic stimulation in cognitive neuroscience: A new synthesis of methodological issues
DOI:10.1016/j.neubiorev.2010.06.005
URL
PMID:20599555
[本文引用: 1]
Transcranial magnetic stimulation (TMS) has become a mainstay of cognitive neuroscience, thus facing new challenges due to its widespread application on behaviorally silent areas. In this review we will summarize the main technical and methodological considerations that are necessary when using TMS in cognitive neuroscience, based on a corpus of studies and technical improvements that has become available in most recent years. Although TMS has been applied only relatively recently on a large scale to the study of higher functions, a range of protocols that elucidate how this technique can be used to investigate a variety of issues is already available, such as single pulse, paired pulse, dual-site, repetitive and theta burst TMS. Finally, we will touch on recent promising approaches that provide powerful new insights about causal interactions among brain regions (i.e., TMS with other neuroimaging techniques) and will enable researchers to enhance the functional resolution of TMS (i.e., state-dependent TMS). We will end by briefly summarizing and discussing the implications of the newest safety guidelines.
Procedural learning: a developmental study of motor sequence learning and probabilistic classification learning in school-aged children
DOI:10.1080/09297049.2015.1058347
URL
PMID:26144545
[本文引用: 2]
In this study, we investigated motor and cognitive procedural learning in typically developing children aged 8-12 years with a serial reaction time (SRT) task and a probabilistic classification learning (PCL) task. The aims were to replicate and extend the results of previous SRT studies, to investigate PCL in school-aged children, to explore the contribution of declarative knowledge to SRT and PCL performance, to explore the strategies used by children in the PCL task via a mathematical model, and to see whether performances obtained in motor and cognitive tasks correlated. The results showed similar learning effects in the three age groups in the SRT and in the first half of the PCL tasks. Participants did not develop explicit knowledge in the SRT task whereas declarative knowledge of the cue-outcome associations correlated with the performances in the second half of the PCL task, suggesting a participation of explicit knowledge after some time of exposure in PCL. An increasing proportion of the optimal strategy use with increasing age was observed in the PCL task. Finally, no correlation appeared between cognitive and motor performance. In conclusion, we extended the hypothesis of age invariance from motor to cognitive procedural learning, which had not been done previously. The ability to adopt more efficient learning strategies with age may rely on the maturation of the fronto-striatal loops. The lack of correlation between performance in the SRT task and the first part of the PCL task suggests dissociable developmental trajectories within the procedural memory system.
The prefrontal cortex and cognitive control
DOI:10.1038/35036228
URL
PMID:11252769
[本文引用: 1]
One of the enduring mysteries of brain function concerns the process of cognitive control. How does complex and seemingly willful behaviour emerge from interactions between millions of neurons? This has long been suspected to depend on the prefrontal cortex--the neocortex at the anterior end of the brain--but now we are beginning to uncover its neural basis. Nearly all intended behaviour is learned and so depends on a cognitive system that can acquire and implement the 'rules of the game' needed to achieve a given goal in a given situation. Studies indicate that the prefrontal cortex is central in this process. It provides an infrastructure for synthesizing a diverse range of information that lays the foundation for the complex forms of behaviour observed in primates.
Intracortical circuits and their interactions in human primary motor cortex
Triple-pulse TMS to study interactions between neural circuits in human cortex
DOI:10.1016/j.brs.2011.01.002
URL
PMID:22032744
[本文引用: 1]
Transcranial magnetic stimulation (TMS) is a widely used brain stimulation technique that allows noninvasive examination of different excitatory and inhibitory circuits at the systems level in the intact human brain. In recent years, considerable knowledge has been accumulated about the physiology of several of these facilitatory and inhibitory processes individually. However, activity in the corresponding neural circuits is not independent of each other. This paper reviews the experiments using triple-pulse TMS that are specifically designed to study interactions between intracortical circuits. These studies have provided evidence for a complex network of interconnected neural circuits within and across cerebral hemispheres. The current knowledge about the functional organization of this network, its pharmacology and functional implications for human motor control are discussed in detail. These findings have clinical relevance because specific interactions between neural circuits may be impaired in neurologic and psychiatric disorders. We conclude that triple-pulse TMS studies will help to integrate and better understand the physiologic processes involved in human motor behavior.
Heterosynaptic modulation of motor cortical plasticity in human
DOI:10.1523/JNEUROSCI.4714-13.2014
URL
PMID:24849363
[本文引用: 1]
Inductions of long-term potentiation (LTP) and depression (LTD) are modulated if they are preceded by a priming protocol, in a manner consistent with metaplasticity. Depotentiation refers to reversal of LTP by a subsequent protocol that has no effect by itself. Paired associative stimulation (PAS) at interstimulus interval of 25 ms (PAS25) and 10 ms (PAS10) produces spike timing-dependent LTP-like and LTD-like effects in human primary motor cortex. Continuous theta burst stimulation (cTBS) with 600 pulses produces an LTD-like effect, whereas cTBS with 150 pulses (cTBS150) has no effect by itself. We investigated whether cortical plasticity induced by PAS can be modulated by heterosynaptic inputs of cTBS150. PAS25 and PAS10 primed and followed by cTBS150 were compared withPAS25 and PAS10 alone. Motor evoked potential (MEP) amplitude, recruitment curve, and intracortical circuits including short-interval intracortical inhibition (SICI), long-interval intracortical inhibition (LICI), intracortical facilitation, and short-latency afferent inhibition were measured before and after the interventions. After PAS25 alone, MEP amplitude increased while intracortical circuits did not change. A priming cTBS150 enhanced the effects of PAS25 with further increase in MEP amplitude and led to reduction in SICI and LICI. PAS25 followed by cTBS150 led to reduced MEP amplitude and increased LICI and SICI. Both priming and following cTBS150 reversed the LTD-like effect produced by PAS10 with little change in intracortical circuits. We conclude that cortical plasticity induced by PAS and cTBS interacts in a heterosynaptic and bidirectional manner. The order of the interventions determines whether the underlying mechanisms are related to metaplasticity or depotentiation.
Two phases of interhemispheric inhibition between motor related cortical areas and the primary motor cortex in human
DOI:10.1093/cercor/bhn201
URL
PMID:19015374
[本文引用: 1]
Interhemispheric inhibition (IHI) refers to the neurophysiological mechanism in which one hemisphere of the brain inhibits the opposite hemisphere. IHI can be studied by transcranial magnetic stimulation using a conditioning-test paradigm. We investigated IHI from 5 motor related cortical areas in the right hemisphere to the left primary motor cortex (M1). These areas are hand and face representations of M1, dorsal premotor cortex, somatosensory cortex, and dorsolateral prefrontal cortex. Test stimulus was delivered to the left M1 and conditioning stimulus (CS) was delivered to one of 5 motor related cortical areas in the right hemisphere. The time course of IHI, effects of different CS intensities and current directions on IHI were tested. Maximum IHI was found at interstimulus intervals of approximately 10 ms (short latency IHI, SIHI) and approximately 50 ms (long latency IHI, LIHI) for the motor related areas tested. LIHI could be elicited over a wide range of CS intensities, whereas SIHI required higher CS intensities. We conclude that there are 2 distinct phases of IHI from motor related cortical areas to the opposite M1 through the corpus callosum, and they are mediated by different neuronal populations.
Influence of measurement order by dominant and nondominant hands on performance of a pursuit-rotor task
DOI:10.2466/PMS.108.3.905-914
URL
PMID:19725325
[本文引用: 1]
This study examined the effect of bilateral transfer and performance variation on 10 trials of a pursuit-rotor task. 95 right-handed healthy men (M age = 21.0 yr., SD = 3.6) were randomly divided into Right-Left (R-L: n = 50) and Left-Right (L-R: n = 45) groups. The former performed the task with the right hand first and the latter group used the left hand first. Target contact times on the pursuit-rotor task improved at about the same rate. Contact times by the left hand after using the right hand were longer than those when the left hand was used first. Although variation of contact times converged with repeated trials, when the left hand was used first, the performance variation remained large for right-handed participants.
A mesoscale connectome of the mouse brain
The assessment and analysis of handedness: The Edinburgh inventory
DOI:10.1016/0028-3932(71)90067-4 URL PMID:5146491 [本文引用: 1]
The role of the dorsolateral prefrontal cortex in implicit procedural learning
DOI:10.1007/bf00230427
URL
PMID:8821387
[本文引用: 2]
We studied the role of the dorsolateral prefrontal cortex in procedural learning. Normal subjects completed several blocks of a serial reaction time task using only one hand without or with concurrent non-invasive repetitive transcranial magnetic stimulation. To disrupt their function transiently, stimulation was applied at low intensity over the supplementary motor area or over the dorsolateral prefrontal cortex contralateral or ipsilateral to the hand used for the test. Stimulation to the contralateral dorsolateral prefrontal cortex markedly impaired procedural implicit learning, as documented by the lack of significant change in response times during the task. Stimulation over the other areas did not interfere with learning. These results support the notion of a critical role of contralateral dorsolateral prefrontal structures in learning of motor sequences.
The neural correlates of motor skill automaticity
DOI:10.1523/JNEUROSCI.3880-04.2005
URL
PMID:15930384
[本文引用: 1]
Acquisition of a new skill is generally associated with a decrease in the need for effortful control over performance, leading to the development of automaticity. Automaticity by definition has been achieved when performance of a primary task is minimally affected by other ongoing tasks. The neural basis of automaticity was examined by testing subjects in a serial reaction time (SRT) task under both single-task and dual-task conditions. The diminishing cost of dual-task performance was used as an index for automaticity. Subjects performed the SRT task during two functional magnetic imaging sessions separated by 3 h of behavioral training over multiple days. Behavioral data showed that, by the end of testing, subjects had automated performance of the SRT task. Before behavioral training, performance of the SRT task concurrently with the secondary task elicited activation in a wide network of frontal and striatal regions, as well as parietal lobe. After extensive behavioral training, dual-task performance showed comparatively less activity in bilateral ventral premotor regions, right middle frontal gyrus, and right caudate body; activity in other prefrontal and striatal regions decreased equally for single-task and dual-task conditions. These data suggest that lateral and dorsolateral prefrontal regions, and their corresponding striatal targets, subserve the executive processes involved in novice dual-task performance. The results also showed that supplementary motor area and putamen/globus pallidus regions showed training-related decreases for sequence conditions but not for random conditions, confirming the role of these regions in the representation of learned motor sequences.
Is there a future for therapeutic use of transcranial magnetic stimulation?
DOI:10.1038/nrn2169
URL
PMID:17565358
[本文引用: 1]
Repetitive transcranial magnetic stimulation (rTMS) has in recent years been used to explore therapeutic opportunities in a bewildering variety of conditions. Although there is good evidence that this technique can modify cortical activity, the rationale for its use in many of the conditions investigated so far is not clear. Here we discuss the effects of rTMS in healthy subjects and how it has been used in a number of neurological conditions. We argue that a better understanding of both the effects of rTMS and the pathological processes underlying the conditions for which it is used will reveal whether rTMS really does offer therapeutic potential and, if so, for which conditions.
The role of the dorsolateral prefrontal cortex during sequence learning is specific for spatial information
DOI:10.1093/cercor/11.7.628
URL
PMID:11415965
[本文引用: 2]
Many studies have implicated the dorsolateral prefrontal cortex in the acquisition of skill, including procedural sequence learning. However, the specific role it performs in sequence learning has remained uncertain. This type of skill has been intensively studied using the serial reaction time task. We used three versions of this task: a standard task where the position of the stimulus cued the response; a non-standard task where the color of the stimulus was related to the correct response; and a combined task where both the color and position simultaneously cued the response. We refer to each of these tasks based upon the cues available for guiding learning as position, color and combined tasks. The combined task usually shows an enhancement of skill acquisition, a result of being driven by two simultaneous and congruent cues. Prior to the performance of each of these tasks the function of the dorsolateral prefrontal cortex was disrupted using repetitive transcranial magnetic stimulation. This completely prevented learning within the position task, while sequence learning occurred to a similar extent in both the color and combined tasks. So, following prefrontal stimulation the expected learning enhancement in the combined task was lost, consistent with only a color cue being available to guide sequence learning in the combined task. Neither of these effects was observed following stimulation at the parietal cortex. Hence the critical role played by the dorsolateral prefrontal cortex in sequence learning is related exclusively to spatial cues. We suggest that the dorsolateral prefrontal cortex operates over the short term to retain and manipulate spatial information to allow cortical and subcortical structures to learn a predictable sequence of actions. Such functions may emerge from the broader role the dorsolateral prefrontal cortex has in spatial working memory. These results argue against the dorsolateral prefrontal cortex constituting part of the neuronal substrate responsible for general aspects of implicit or explicit sequence learning.
Using transcranial magnetic stimulation methods to probe connectivity between motor areas of the brain
DOI:10.1016/j.humov.2010.07.007
URL
PMID:21056490
[本文引用: 1]
Transcranial magnetic stimulation is increasingly used as a tool to explore cortical motor function in healthy subjects and in patients with neurological disease or injury. This review describes a &quot;twin coil&quot; TMS approach that allows investigation of time related changes in functional connectivity between primary motor cortex and other areas in preparation for a forthcoming movement. Investigations into premotor-motor interactions show that these are specific to the type of task that is performed as well as the muscles used to control the movement, allowing us to monitor information flow within motor networks with millisecond time resolution.
The prefrontal cortex shows context-specific changes in effective connectivity to motor or visual cortex during the selection of action or colour
DOI:10.1093/cercor/bhh111
URL
PMID:15238443
[本文引用: 1]
The role of the prefrontal cortex remains controversial. Neuroimaging studies support modality-specific and process-specific functions related to working memory and attention. Its role may also be defined by changes in its influence over other brain regions including sensory and motor cortex. We used functional magnetic imaging (fMRI) to study the free selection of actions and colours. Control conditions used externally specified actions and colours. The prefrontal cortex was activated during free selection, regardless of modality, in contrast to modality-specific activations outside prefrontal cortex. Structural equation modelling (SEM) of fMRI data was used to test the hypothesis that although the same regions of prefrontal cortex may be active in tasks within different domains, there is task-dependent effective connectivity between prefrontal cortex and non-prefrontal cortex. The SEM included high-order interactions between modality, selection and regional activity. There was greater coupling between prefrontal cortex and motor cortex during free selection and action tasks, and between prefrontal cortex and visual cortex during free selection of colours. The results suggest that the functions of the prefrontal cortex may be defined not only by selection-specific rather than modality-specific processes, but also by changing patterns of effective connectivity from prefrontal cortex to motor and sensory cortices.
Transition of brain activation from frontal to parietal areas in visuomotor sequence learning
We studied the neural correlates of visuomotor sequence learning using functional magnetic resonance imaging (fMRI). In the test condition, subjects learned, by trial and error, the correct order of pressing two buttons consecutively for 10 pairs of buttons (2 x 10 task); in the control condition, they pressed buttons in any order. Comparison between the test condition and the control condition revealed four brain areas specifically related to learning: the dorsolateral prefrontal cortex (DLPFC), the presupplementary motor area (pre-SMA), the precuneus, and the intraparietal sulcus (IPS). We found that the time course of activation during learning was different between these areas. To normalize the individual differences in the speed of learning, we classified the performance of each subject into three learning stages: early, intermediate, and advanced stages. Both the relative increase of signal intensity and the number of activated pixels within the four areas showed significant changes across the learning stages, with different time courses. The two frontal areas, DLPFC and pre-SMA, were activated in the earlier stages of learning, whereas the two parietal areas, precuneus and IPS, were activated in the later stages. Specifically, DLPFC, pre-SMA, precuneus, and IPS were most highly activated in the early stage, in both the early and intermediate stages, in the intermediate stage, and in both the intermediate and advanced stages, respectively. The results suggest that the acquisition of visuomotor sequences requires frontal activation, whereas the retrieval of visuomotor sequences requires parietal activation, which might reflect the transition from the declarative stage to the procedural stage.
An FMRI study of the role of the medial temporal lobe in implicit and explicit sequence learning
DOI:10.1016/s0896-6273(03)00123-5
URL
PMID:12670429
[本文引用: 1]
fMRI was used to investigate the neural substrates supporting implicit and explicit sequence learning, focusing especially upon the role of the medial temporal lobe. Participants performed a serial reaction time task (SRTT). For implicit learning, they were naive about a repeating pattern, whereas for explicit learning, participants memorized another repeating sequence. fMRI analyses comparing repeating versus random sequence blocks demonstrated activation of frontal, parietal, cingulate, and striatal regions implicated in previous SRTT studies. Importantly, mediotemporal lobe regions were active in both explicit and implicit SRTT learning. Moreover, the results provide evidence of a role for the hippocampus and related cortices in the formation of higher order associations under both implicit and explicit learning conditions, regardless of conscious awareness of sequence knowledge.
Neural correlates of encoding and expression in implicit sequence learning
DOI:10.1007/s00221-005-2284-z
URL
PMID:15965762
[本文引用: 1]
In the domain of motor learning it has been difficult to separate the neural substrate of encoding from that of change in performance. Consequently, it has not been clear whether motor effector areas participate in learning or merely modulate changes in performance. Here, using a variant of the serial reaction time task that dissociated these two factors, we report that encoding during procedural motor learning does engage cortical motor areas and can be characterized by distinct early and late encoding phases. The highest correlation between activation and subsequent changes in motor performance was seen in the motor cortex during early encoding, and in the basal ganglia during the late encoding phase. Our results show that rapid encoding during procedural motor learning involves several distinct processes, and is represented primarily within motor system structures.
Neural correlates of motor memory consolidation
DOI:10.1126/science.277.5327.821
URL
PMID:9242612
[本文引用: 1]
Computational studies suggest that acquisition of a motor skill involves learning an internal model of the dynamics of the task, which enables the brain to predict and compensate for mechanical behavior. During the hours that follow completion of practice, representation of the internal model gradually changes, becoming less fragile with respect to behavioral interference. Here, functional imaging of the brain demonstrates that within 6 hours after completion of practice, while performance remains unchanged, the brain engages new regions to perform the task; there is a shift from prefrontal regions of the cortex to the premotor, posterior parietal, and cerebellar cortex structures. This shift is specific to recall of an established motor skill and suggests that with the passage of time, there is a change in the neural representation of the internal model and that this change may underlie its increased functional stability.
Declarative and nondeclarative memory: multiple brain systems supporting learning and memory
DOI:10.1162/jocn.1992.4.3.232
URL
PMID:23964880
[本文引用: 1]
The topic of multiple forms of memory is considered from a biological point of view. Fact-and-event (declarative, explicit) memory is contrasted with a collection of non conscious (non-declarative, implicit) memory abilities including skills and habits, priming, and simple conditioning. Recent evidence is reviewed indicating that declarative and non declarative forms of memory have different operating characteristics and depend on separate brain systems. A brain-systems framework for understanding memory phenomena is developed in light of lesion studies involving rats, monkeys, and humans, as well as recent studies with normal humans using the divided visual field technique, event-related potentials, and positron emission tomography (PET).
Shifts in connectivity during procedural learning after motor cortex stimulation: A combined transcranial magnetic stimulation/functional magnetic resonance imaging study
DOI:10.1016/j.cortex.2015.10.004
URL
PMID:26673946
[本文引用: 1]
Inhibitory transcranial magnetic stimulation (TMS), of which continuous theta burst stimulation (cTBS) is a common form, has been used to inhibit cortical areas during investigations of their function. cTBS applied to the primary motor area (M1) depresses motor output excitability via a local effect and impairs procedural motor learning. This could be due to an effect on M1 itself and/or to changes in its connectivity with other nodes in the learning network. To investigate this issue, we used functional magnetic resonance imaging to measure changes in brain activation and connectivity during implicit procedural learning after real and sham cTBS of M1. Compared to sham, real cTBS impaired motor sequence learning, but caused no local or distant changes in brain activation. Rather, it reduced functional connectivity between motor (M1, dorsal premotor &amp; supplementary motor areas) and visual (superior &amp; inferior occipital gyri) areas. It also increased connectivity between frontal associative (superior &amp; inferior frontal gyri), cingulate (dorsal &amp; middle cingulate), and temporal areas. This potentially compensatory shift in coupling, from a motor-based learning network to an associative learning network accounts for the behavioral effects of cTBS of M1. The findings suggest that the inhibitory TMS affects behavior via relatively subtle and distributed effects on connectivity within networks, rather than by taking the stimulated area &quot;offline&quot;.
Temporary occlusion of associative motor cortical plasticity by prior dynamic motor training
DOI:10.1093/cercor/bhi116
URL
PMID:15930370
[本文引用: 1]
A novel Hebbian stimulation paradigm was employed to examine physiological correlates of motor memory formation in humans. Repetitive pairing of median nerve stimulation with transcranial magnetic stimulation over the contralateral motor cortex (paired associative stimulation, PAS) may decrease human motor cortical excitability at interstimulus intervals of 10 ms (PAS10) or increase excitability at 25 ms (PAS25). The properties of this plasticity have previously been shown to resemble associative timing-dependent long-term depression (LTD) and long-term potentiation (LTP) as established in vitro. Immediately after training a novel dynamic motor task, the capacity of the motor cortex to undergo plasticity in response to PAS25 was abolished. PAS10-induced plasticity remained unchanged. When retested after 6 h, PAS25-induced plasticity recovered to baseline levels. After training, normal PAS25-induced plasticity was observed in the contralateral training-naive motor cortex. Motor training did not reduce the efficacy of PAS25 to enhance cortical excitability when PAS10 was interspersed between the training and application of the PAS25 protocol. This indicated that the mechanism supporting PAS25-induced plasticity had remained intact immediately after training. Behavioral evidence was obtained for continued optimization of force generation at a time when PAS25-induced plasticity was blocked in the training motor cortex. Application of the PAS protocols after motor training did not prevent the consolidation of motor skills evident as performance gains at later retesting. The results are consistent with a concept of temporary suppression of associative cortical plasticity by neuronal mechanisms involved in motor training. Although it remains an open question exactly which element of motor training was responsible for this effect, our findings may link dynamic properties of LTP formation, as established in animal experiments, with human motor memory formation and possibly dynamic motor learning.
The time course of changes during motor sequence learning: A whole-brain fMRI study
DOI:10.1006/nimg.1998.0349
URL
PMID:9698575
[本文引用: 3]
There is a discrepancy between the results of imaging studies in which subjects learn motor sequences. Some experiments have shown decreases in the activation of some areas as learning increased, whereas others have reported learning-related increases as learning progressed. We have exploited fMRI to measure changes in blood oxygen leve-dependent (BOLD) signal throughout the course of learning. T2*-weighted echo-planar images were acquired over the whole brain for 40 min while the subjects learned a sequence eight moves long by trial and error. The movements were visually paced every 3.2 s and visual feedback was provided to the subjects. A baseline period followed each activation period. The effect due to the experimental conditions was modeled using a square-wave function, time locked to their occurrence. Changes over time in the difference between activation and baseline signal were modeled using a set of polynomial basis functions. This allowed us to take into account linear as well as nonlinear changes over time. Low-frequency changes over time common to both activation and baseline conditions (and thus not learning related) were modeled and removed. Linear and nonlinear changes of BOLD signal over time were found in prefrontal, premotor, and parietal cortex and in neostriatal and cerebellar areas. Single-unit recordings in nonhuman primates during the learning of motor tasks have clearly shown increased activity early in learning, followed by a decrease as learning progressed. Both phenomena can be observed at the population level in the present study.
Neural mechanisms underlying the changes in ipsilateral primary motor cortex excitability during unilateral rhythmic muscle contraction
DOI:10.1016/j.bbr.2012.10.053
URL
PMID:23174210
[本文引用: 1]
The aim of this study was to investigate the neural mechanisms underlying the changes in the ipsilateral primary motor cortex (ipsi-M1) excitability induced during the unilateral rhythmic muscle contraction of the first dorsal interosseous (FDI) (rhythmic contraction) muscle with three different frequencies of auditory cues (1, 2, and 3 Hz). The effect of different frequencies of unilateral rhythmic contraction on changes in the ipsi-M1 excitability was assessed using a single-pulse transcranial magnetic stimulation (TMS) technique when subjects were performing the unilateral rhythmic contractions according to each auditory cue frequency. After that, the changes in short intracortical inhibition (SICI)/facilitation (ICF), long intracortical inhibition (LICI) within the ipsi-M1, and interhemispheric inhibition (IHI), as well as dorsal premotor cortex to M1 (PMd-M1), and dorsolateral prefrontal cortex to M1 (DLPFC-M1) connectivity from the contralateral hemisphere to the ipsi-M1 were assessed using paired-pulse TMS techniques. The motor evoked potentials (MEP) induced in the right FDI were recorded. In the results, the ipsi-M1 excitability induced in response to single-pulse TMS was significantly decreased in the 2 Hz conditions, compared with the 1Hz and 3Hz conditions. Furthermore, PMd-M1 connectivity and LICI were significantly modulated depending on the frequency of the unilateral rhythmic contraction. In contrast, the changes in the SICI, ICF, IHI, and DLPFC-M1 were not directly associated with the rhythm frequency. These results suggest that PMd-M1 connectivity and LICI within the ipsi-M1 are likely to preferentially operate to modulate ipsi-M1 excitability during the performance of unilateral rhythmic contraction with different frequencies.
Modulation of motor cortical excitability by electrical stimulation over the cerebellum in man
DOI:10.1007/s004240000402
URL
PMID:11205062
[本文引用: 1]
An alternatively spliced isoform of the P2X1 receptor (P2X1a) was cloned from rat mesenteric artery. The spliced isoform does not have the 27 amino acids that are in the middle of the putative extracellular loop domain of the P2X1 original subunit (P2X1). Reverse transcriptase polymerase chain reaction (RT-PCR) revealed co-localization of P2X1a mRNA and P2X1 mRNA in vascular and other smooth muscle tissues and heart, but not in the spinal cord. In HEK293 cells transfected with P2X1 cDNA, ATP (1 microM) evoked an inward current which strongly desensitized, and an intense signal for GFP (green fluorescent protein) fused with P2X1 was detected at the membrane. Neither of these results was obtained in HEK293 cells expressing P2X1a alone. The fluorescent GFP signal was detected at the membrane when GFP-fused P2X1a was co-expressed with P2X1, and no significant difference in the ATP-activated current was noted between cells expressing P2X1 and those coexpressing P2X1 and P2X1a. These results indicate that the 27-amino-acid sequence (175-201) is important for protein trafficking to the membrane and for the formation of a functional P2X1 receptor. Our results also show that P2X1a is transported to the membrane when P2X1a is co-expressed with P2X1, although the co-expression of P2X1a does not modify the channel's current properties.
The geometric structure of the brain fiber pathways
DOI:10.1126/science.1215280
URL
PMID:22461612
[本文引用: 2]
The structure of the brain as a product of morphogenesis is difficult to reconcile with the observed complexity of cerebral connectivity. We therefore analyzed relationships of adjacency and crossing between cerebral fiber pathways in four nonhuman primate species and in humans by using diffusion magnetic resonance imaging. The cerebral fiber pathways formed a rectilinear three-dimensional grid continuous with the three principal axes of development. Cortico-cortical pathways formed parallel sheets of interwoven paths in the longitudinal and medio-lateral axes, in which major pathways were local condensations. Cross-species homology was strong and showed emergence of complex gyral connectivity by continuous elaboration of this grid structure. This architecture naturally supports functional spatio-temporal coherence, developmental path-finding, and incremental rewiring with correlated adaptation of structure and function in cerebral plasticity and evolution.
A neuropsychological theory of motor skill learning
DOI:10.1037/0033-295x.105.3.558
URL
PMID:9697430
[本文引用: 2]
This article describes a neuropsychological theory of motor skill learning that is based on the idea that learning grows directly out of motor control processes. Three motor control processes may be tuned to specific tasks, thereby improving performance: selecting spatial targets for movement, sequencing these targets, and transforming them into muscle commands. These processes operate outside of awareness. A 4th, conscious process can improve performance in either of 2 ways: by selecting more effective goals of what should be changed in the environment or by selecting and sequencing spatial targets. The theory accounts for patterns of impairment of motor skill learning in patient populations and for learning-related changes in activity in functional imaging studies. It also makes a number of predictions about the purely cognitive, including accounts of mental practice, the representation of motor skill, and the interaction of conscious and unconscious processes in motor skill learning.
Direct comparison of neural systems mediating conscious and unconscious skill learning
DOI:10.1152/jn.2002.88.3.1451
URL
PMID:12205165
[本文引用: 1]
Procedural learning, such as perceptual-motor sequence learning, has been suggested to be an obligatory consequence of practiced performance and to reflect adaptive plasticity in the neural systems mediating performance. Prior neuroimaging studies, however, have found that sequence learning accompanied with awareness (declarative learning) of the sequence activates entirely different brain regions than learning without awareness of the sequence (procedural learning). Functional neuroimaging was used to assess whether declarative sequence learning prevents procedural learning in the brain. Awareness of the sequence was controlled by changing the color of the stimuli to match or differ from the color used for random sequences. This allowed direct comparison of brain activation associated with procedural and declarative memory for an identical sequence. Activation occurred in a common neural network whether initial learning had occurred with or without awareness of the sequence, and whether subjects were aware or not aware of the sequence during performance. There was widespread additional activation associated with awareness of the sequence. This supports the view that some types of unconscious procedural learning occurs in the brain whether or not it is accompanied by conscious declarative knowledge.
Implicit sequence learning of emotional stimulation and semantic stimulation (Unpublished master dissertation)
情绪刺激和语义刺激的内隐序列学习研究(硕士学位论文)
Neurophysiological mechanisms of motor learning
动作技能学习神经生理机制研究
Embodied mechanisms of implicit sequence learning consciousness
DOI:10.3724/SP.J.1042.2016.00203
URL
[本文引用: 1]
There are three theories of implicit sequence learning consciousness: global workspace theory, neural plasticity theory and extraneous stimulus theory. All of them ignore body sense which is the key factor of consciousness. Therefore it is difficult for them to reveal the fundamental cause of consciousness. Embodied consciousness theory and its studies showed that motor/emotional mirror neuron system and their interaction with self and control systems are the source of primary/advanced consciousness. However, they did not involve the rule consciousness of implicit sequence learning which is very important to human study and cognition. In essence, previous researches on implicit sequence learning were close to reveal that the mechanism of implicit sequence learning is just sense/motor embodied learning, that its consciousness mechanism is likely to be motor/emotional embodied consciousness, and that the consciousness processing brain areas of implicit sequence learning mostly overlap the embodied consciousness brain areas. Future research should use Granger causality brain network technology to prove the embodied origin of implicit sequence learning consciousness, to examine the embodied basis of the three kinds of consciousness theories, and to explore the embodied mechanism of factors influencing consciousness.
内隐序列学习意识的具身机制
The effects of motor skill level and somatosensory input on motor imagery: An fMRI study on basketball free shot
DOI:10.3724/SP.J.1041.2017.00307 URL [本文引用: 1]
运动技能水平与躯体感觉输入对运动表象的影响
TMS and drugs
DOI:10.1016/j.clinph.2004.03.006
URL
PMID:15261850
[本文引用: 1]
The application of a single dose of a CNS active drug with a well-defined mode of action on a neurotransmitter or neuromodulator system may be used for testing pharmaco-physiological properties of transcranial magnetic stimulation (TMS) measures of cortical excitability. Conversely, a physiologically well-defined single TMS measure of cortical excitability may be used as a biological marker of acute drug effects at the systems level of the cerebral cortex. An array of defined TMS measures may be used to study the pattern of effects of a drug with unknown or multiple modes of action. Acute drug effects may be rather different from chronic drug effects. These differences can also be studied by TMS measures. Finally, TMS or repetitive TMS by themselves may induce changes in endogenous neurotransmitters or neuromodulators. All these possible interactions are the focus of this in-depth review on TMS and drugs.
TMS and drugs revisited 2014
DOI:10.1016/j.clinph.2014.08.028
URL
PMID:25534482
[本文引用: 1]
The combination of pharmacology and transcranial magnetic stimulation to study the effects of drugs on TMS-evoked EMG responses (pharmaco-TMS-EMG) has considerably improved our understanding of the effects of TMS on the human brain. Ten years have elapsed since an influential review on this topic has been published in this journal (Ziemann, 2004). Since then, several major developments have taken place: TMS has been combined with EEG to measure TMS evoked responses directly from brain activity rather than by motor evoked potentials in a muscle, and pharmacological characterization of the TMS-evoked EEG potentials, although still in its infancy, has started (pharmaco-TMS-EEG). Furthermore, the knowledge from pharmaco-TMS-EMG that has been primarily obtained in healthy subjects is now applied to clinical settings, for instance, to monitor or even predict clinical drug responses in neurological or psychiatric patients. Finally, pharmaco-TMS-EMG has been applied to understand the effects of CNS active drugs on non-invasive brain stimulation induced long-term potentiation-like and long-term depression-like plasticity. This is a new field that may help to develop rationales of pharmacological treatment for enhancement of recovery and re-learning after CNS lesions. This up-dated review will highlight important knowledge and recent advances in the contribution of pharmaco-TMS-EMG and pharmaco-TMS-EEG to our understanding of normal and dysfunctional excitability, connectivity and plasticity of the human brain.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}