

1 引言
急性应激会影响个体对威胁刺激的注意偏向(Jiang et al., 2017; Sänger, Bechtold, Schoofs, Blaszkewicz, & Wascher, 2014; Weymar, Schwabe, Löw, & Hamm, 2012), 但是这种影响的机制不清楚, 即急性应激是增强了个体早期对威胁刺激的注意定向还是损害了晚期对威胁刺激的注意解除还不清楚, 因此本研究探讨急性应激影响个体对威胁刺激注意偏向的认知机制。
对威胁刺激的注意偏向是指个体对威胁刺激与中性刺激的注意分配不同, 个体对威胁刺激存在偏向。其主要表现为对威胁刺激的注意定向加速和对威胁刺激的注意解除困难(Koster, Crombez, Verschuere, van Damme, & Wiersema, 2006; Zhang, Luo, Zhao, Chen, & Li, 2014)。对威胁刺激的注意定向加速是指相对于中性刺激, 个体能够更快地将注意指向威胁刺激或具有威胁性的位置(Carlson & Reinke, 2008); 对威胁刺激的注意解除困难是指与中性刺激相比, 个体更难停止对威胁刺激或者威胁性位置的注意加工(Fox, Russo, Bowles, & Dutton, 2001)。点探测任务经常被用于测量个体对威胁刺激的注意定向和注意解除。在点探测任务中, 以威胁刺激和中性刺激组成的注意分配条件(威胁-中性刺激对)和非注意分配条件(威胁-威胁刺激对、中性-中性刺激对)作为线索, 之后一个中性目标随机出现在威胁或中性线索的位置, 个体对目标进行反应。在注意分配条件中, 目标出现在威胁线索位置的条件称为一致条件, 出现在中性线索位置称为不一致条件; 在非注意分配条件, 由于线索属性相同, 个体不会产生注意偏向(Koster, Crombez, Verschuere, & de Houwer, 2004)。注意定向加速是指个体在非注意分配条件作为线索时对目标刺激的反应慢于一致条件; 而注意解除困难则是指个体在不一致条件作为线索时对目标刺激的反应慢于非注意分配条件(Salemink, van den Hout, & Kindt, 2007)。
已有研究使用事件相关电位(Event-Related Potentials, ERP)技术发现, 个体对威胁刺激注意定向和注意解除分别与N2pc (N2-posteior- contralateral)和SPCN (sustained posterior contralateral negativity)相关(Holmes, Bradley, Kragh Nielsen, & Mogg, 2009; Holmes, Mogg, de Fockert, Nielsen, & Bradley, 2014; Kappenman, Farrens, Luck, & Proudfit, 2014; Kappenman, MacNamara, & Proudfit, 2015)。N2pc是视觉目标对侧视野与目标同侧视野(如:威胁刺激在右侧, 则左半球电极点为对侧, 右半球电极点为同侧)电极点的波幅差异波, 分布在大脑后部, 在P7/P8和PO7/PO8电极周围波幅最大, 一般出现在目标刺激呈现之后约180~300 ms, 其波幅反映了个体对目标刺激的注意选择(Holmes et al., 2009; Holmes et al., 2014; Kappenman et al., 2014; Kappenman et al., 2015)。研究发现, 威胁刺激会诱发更负的N2pc, 表明威胁刺激更快地捕获了注意(Holmes et al., 2009; Holmes et al., 2014; Kappenman et al., 2014; Kappenman et al., 2015)。这种对威胁刺激的注意定向可能与以杏仁核为核心的杏仁核-前扣带回网络紧密相关(Carlson et al., 2012; Carlson & Reinke, 2008)。SPCN同N2pc一样, 也是大脑后部对侧更负的偏侧事件相关电位, 一般出现在目标刺激呈现300 ms 以后。SPCN 的波幅反映的是个体对目标刺激的注意维持过程(Holmes et al., 2009; Holmes et al., 2014)。研究也发现威胁刺激会诱发更大的SPCN, 表明个体对威胁刺激存在注意维持, 即个体更难将注意从威胁刺激中解除出来(Holmes et al., 2009; Holmes et al., 2014; Meconi, Luria, & Sessa, 2014; Sessa, Luria, Gotler, Jolicœur, & Dell'acqua, 2011)。这种对威胁刺激的注意解除可能与包括背外侧前额叶与外侧后顶叶皮层的额-顶网络有关(Fecteau & Munoz, 2006; Gottlieb, 2007)。
急性应激是指无法预期和控制的环境要求超出个体的自然调节能力时, 个体做出的非特异性反应(Koolhaas et al., 2011)。在个体主观感受上, 急性应激会使负面情绪(如:焦虑等)显著增加; 在生理系统上, 急性应激会激活个体的自主神经系统, 诱发儿茶酚胺(如:去甲肾上腺素)的分泌, 并与下丘脑-垂体-肾上腺轴(Hypothalamic-Pituitary-Adrenal axis, HPA 轴)激活导致释放的糖皮质激素(如:皮质醇)结合从而影响个体的神经和生理活动(Ulrich-Lai & Herman, 2009)。急性应激导致的神经和生理活动变化会对杏仁核-前扣带回网络和额-顶网络等脑区域产生特异性影响, 这两个网络与个体对威胁刺激的注意偏向紧密相关(Arnsten, 2009; Hermans et al., 2011)。一方面, 急性应激会增强杏仁核-前扣带回网络的活性(Cousijn et al., 2010), 促进个体对威胁刺激的检测(Dȩbiec & LeDoux, 2006)。Cousijn等人研究发现急性应激增强了杏仁核的阶段性反应(Cousijn et al., 2010)。Rued等人的行为学研究发现, 急性应激状态下, 个体对威胁刺激的反应速度更快但准确率更低, 个体对威胁刺激的行为反应优势明显(Rued, Hilmert, Strahm, & Thomas, 2019)。同时在ERP研究中也发现急性应激会增强个体对威胁刺激的早期感觉输入, 表现为早期与注意相关的N1成分增大(Löw, Weymar, & Hamm, 2015; Shackman, Maxwell, McMenamin, Greischar, & Davidson, 2011)。个体对威胁刺激的注意定向与杏仁核-前扣带回网络紧密相关(Carlson et al., 2012; Carlson & Reinke, 2008), 急性应激会增加该网络的活动。据此,我们推测急性应激会增强个体对威胁刺激的注意定向。另一方面, 急性应激会降低额-顶网络的活性(Arnsten, 2009; Qin, Hermans, van Marle, Luo, & Fernández, 2009), 削弱个体对注意控制资源的分配(Meconi et al., 2014)。Qin等人研究发现急性应激显著降低了大脑区域中前额叶皮层、顶叶皮层的激活(Qin et al., 2009)。同时, 与急性应激相关疾病(如焦虑症、抑郁症等)的ERP研究也发现了类似的结果。Meconi等人发现与低焦虑个体相比, 不可信任面孔(威胁刺激)诱发高焦虑个体产生了更负的SPCN波幅, 表明高焦虑个体比低焦虑个体更难将注意从不可信任面孔上解除出来(Meconi et al., 2014)。个体对威胁刺激的注意解除与额-顶网络有关(Fecteau & Munoz, 2006; Gottlieb, 2007), 急性应激会损害该网络的活动。因此,我们推测急性应激会损害个体对威胁刺激的注意解除。
综上, 急性应激可能会对注意偏向的不同表现产生不同的影响。即急性应激可能会增强个体对威胁刺激的注意定向, 损害个体对威胁刺激的注意解除。基于此, 本研究采用社会评估冷压任务(socially evaluated cold pressor test, SECPT)诱发个体的应激反应后进行点探测任务, 并结合事件相关电位技术, 研究急性应激影响个体对威胁刺激注意偏向的认知机制。本研究假设, 急性应激会增强个体对威胁刺激的注意定向, 损害个体对威胁刺激的注意解除。如果急性应激增强个体对威胁刺激的注意定向, 则在行为上应激组比控制组表现出更快的注意定向加速, 非注意分配条件与一致条件的反应时之差大于控制组; 在ERP结果上, 威胁刺激会诱发应激组个体产生更负的N2pc。如果急性应激损害个体对威胁刺激的注意解除, 则应激组比控制组在行为上会表现出更慢的注意解除, 不一致条件与非注意分配条件的反应时之差大于控制组; 在ERP结果上, 威胁刺激会诱发应激组个体产生更负的SPCN。
2 方法
2.1 被试
考虑到女性生理周期对应激反应的影响(Fernández et al., 2003)且无法招募到处于不同生理周期的满足实验条件设置数量的女性被试, 所以本研究将采用方便取样的方法选取44名男性成年被试(其中, 应激组24名, 控制组20名), 年龄范围在18~21岁之间, 右利手, 被试自述无躯体疾病及精神障碍, 参与实验前的1个月内未服用过类固醇之类的药物。裸视或矫正视力正常。实验得到当地伦理委员会的批准。实验遵循自愿参与的原则, 在实验之前签署知情同意书, 实验结束后给予一定的报酬。在ERP数据分析中, 对平均波幅在1.25微伏左右, 被试数量在20名左右时, 平均叠加的试次数在100个以上, 结果才稳定可靠(Boudewyn, Luck, Farrens, & Kappenman, 2018)。因此, 数据分析中有8名被试由于脑电伪迹检测后每个条件可供平均叠加的试次数小于100个而被排除, 最后对36名被试(应激组和控制组各18人)的数据进行分析。
2.2 程序
2.2.1 实验刺激与问卷
所有的实验刺激均通过屏幕背景为黑色的17寸显示屏(分辨率为1024×768, 刷新率为60 Hz)呈现。情绪面孔均选自中国情绪面孔图片系统(CAFPS) (Wang & Luo, 2005)的中性和恐惧面孔, 均为黑白图片, 图片大小均为7°×9°。实验总共包含100张情绪面孔, 其中恐惧50张, 中性面孔50张, 男女各半, 每张面孔图片的身份不同。
状态-特质焦虑问卷(State-Trait Anxiety Inventory, STAI)由Spielberger等(1970)编制, 含状态焦虑(S-AI)和特质焦虑(T-AI)两个分量表, 每个分量表有20个项目, 采用4级评分, 从“完全没有”到“非常明显”, 正性情绪项目和负性情绪项目各有一半, 正性情绪项目均为反向计分, 项目累加得分越高, 焦虑程度越严重(Spielberger, 1970)。本研究中该量表的 Cronbach’s α系数为0.89, 状态焦虑分量表和特质焦虑分量表的Cronbach’s α系数分别为0.93、0.79。
2.2.2 实验程序与实验任务
实验采用2(组别:应激组/控制组) × 3(试次条件:一致/不一致/非注意分配)的混合实验设计, 其中组别为被试间因素。为避免被试自身皮质醇的节律对实验结果的影响, 所有的研究都安排在13:30~ 18:30之间进行(Luo et al., 2018)。实验开始前四天电话联系, 告知被试, 实验前72小时之内不饮酒, 实验前24时之内不抽烟, 实验当天不做剧烈运动, 实验前2小时内, 除饮水外, 不能吃任何食物。具体实验流程是(如图1(a)所示):被试到达实验室, 在独立房间阅读并签署知情同意书后静坐休息5分钟, 之后填写第一次状态-特质焦虑量表并收集唾液样本S1 (-85 min), 使用德国SARSTEDT唾液采集管收集唾液, 要求被试将棉条置于口中旋转2分钟, 不能咀嚼; 继续静坐休息20分钟后填写第二次状态焦虑问卷并收集唾液样本S2 (-55 min)。准备结束后被试被随机分配到应激组或者控制组, 并进行SECPT任务或者对应的控制任务(-5 min), 任务结束后填写第三次状态焦虑问卷并收集唾液样本S3 (0 min)。然后被试开始进行点探测任务并同步记录行为和脑电数据, 点探测任务结束后填写第四次状态焦虑问卷并收集唾液样本S4 (40 min), 之后被试休息20分钟后填写第五次状态焦虑问卷并收集唾液样本S5 (70 min), 完成之后离开实验室。
图1
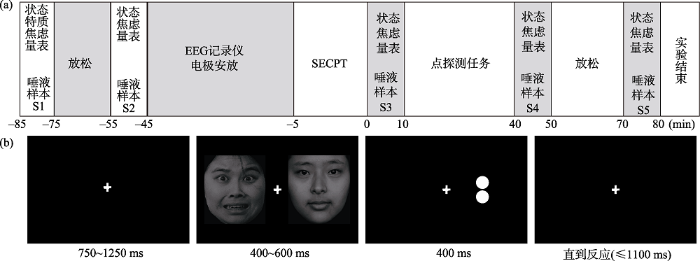
社会评估冷压任务:是由Schwabe等人提出的(Schwabe, Haddad, & Schachinger, 2008; Schwabe & Schächinger, 2018)。在应激条件下, 被试被要求一直注视摄像机, 2名主试穿着医用服站在被试前面以严肃的表情持续评估被试的表现, 与摄像机保持一定的距离, 以便主试、被试和摄像机之间形成一个等边三角形, 保证被试在注视摄像机的同时可以从眼角处看见主试, 之后让被试将脚(含脚腕)浸入冰水(0~2℃), 将脚而不是手浸入冰水是为了防止手部过于冰冷而影响了被试的按键反应, 被试不知道浸泡的具体时间, 但可以随时将脚移出冰水, 在冰水中的时间最长不超过3分钟, 主试会在3分钟时提醒被试将脚移出冰水, 在应激任务进行中, 主试不能说话, 避免任何形式的积极强化(如:微笑、点头等)。在控制条件下, 主试穿着便服站在被试前面以中性的表情持续评估被试的表现, 无摄像机,要求被试将脚(含脚腕)浸入温水(36~38℃)中持续时间至少3分钟。
点探测任务:采用修订版本的点探测任务来测量注意偏向。在每个试次中(见图1(b)), 首先屏幕中央在750~1250 ms内随机呈现一个“+”注视点, 之后在400~600 ms内随机呈现一组由恐惧面孔和中性面孔随机组合的情绪面孔对, 情绪面孔对包括混合对(恐惧面孔-中性面孔/中性面孔-恐惧面孔)、中性对(中性面孔-中性面孔)和恐惧对(恐惧面孔-恐惧面孔)三种类型, 其中面孔对均为同性别匹配, 每张面孔呈现的位置随机且呈现次数不超过9次。然后在注视点的左侧或右侧随机呈现400 ms的目标刺激, 目标刺激为水平或垂直方向组合的两个圆点, 圆点的大小均为0.5° ×0.5°, 目标刺激界面消失后, 被试需要在保证正确率的同时在1100 ms内按k或l键判断圆点的方向(垂直或水平), 按键在被试间平衡, 若1100 ms内无按键反应则视为反应错误并自动进入下一试次。混合对有360个试次, 中性对与恐惧对各60个试次, 总共480个试次。其中, 混合对360个试次中180个试次为目标替换恐惧面孔呈现的位置(一致条件)和180个试次为目标替换中性面孔呈现的位置(不一致条件)。被试的眼睛与屏幕正中保持水平并距离60 cm左右。采用E-prime软件进行刺激呈现和行为学数据收集。
2.3 数据记录与分析
2.3.1 唾液储存与化验
采集后的唾液样本冷藏于-20℃的冰柜中。唾液化验前, 将唾液样本溶解后在3000 rmp转速下离心10分钟。采用电化学发光免疫分析法测定唾液皮质醇浓度(Cobas e 601, Roche Diagnostics, Numbrecht, Germany)。敏感性为1.5 nmol/L (下限)。
2.3.2 行为学数据分析
在点探测任务中, 分别对每个被试的行为学数据进行如下的数据处理:(1)剔除反应错误的试次; (2)剔除反应时小于200 ms和大于1000 ms的试次; (3)剔除反应时在3个标准差之外的试次。经过以上数据处理步骤后, 剔除试次的比率范围在0.63% ~ 13.75%之间, 平均比率为4.91%。对纳入分析的被试, 分别计算注意定向和注意解除的分数后, 进行2(组别:应激组/控制组) × 2(注意偏向:注意定向/注意解除)的重复测量方差分析。注意定向是非注意分配条件的平均反应时减去一致条件下的平均反应时; 注意解除是不一致条件下的平均反应时减去非注意分配条件的平均反应时(Salemink et al., 2007)。
2.3.3 EEG记录与数据分析
采用NeuroScan的ERP记录系统, 按照国际10-20系统扩展的64导电极帽记录EEG。在线记录时以左侧乳突为参考电极。垂直眼电(VEOG)与水平眼电(HEOG)均为双极记录, VEOG电极分别置于左眼眶上和眶下正中, HEOG置于左右外眼角外侧。接地点位于头皮前中部FPz和Fz电极点连线的中点。采样频率为1000 Hz, 滤波带通为0.05~100 Hz, 电极与头皮之间的阻抗均小于5 kΩ。
使用Matlab (R2013b)、EEGLAB (v14.1.2)、ERPLAB (v7.0.0)对连续EEG数据进行离线分析。以双侧乳突平均作为参考, 分别进行高通滤波(0.1 Hz, 12 dB/octave)和低通滤波(30 Hz, 24 dB/octave), 以情绪面孔对刺激出现为标记, 刺激出现前200 ms到刺激出现后600 ms为时间窗口对连续数据进行分段, 并选取-200~0 ms脑电作为基线。分别使用ERPLAB (v7.0.0)中Moving Window Peak-to-Peak (Width: 200 ms; Step Size: 100 ms)和Simple Voltage Threshold自动矫正眨眼伪迹, 运动伪迹剔除阈限为± 100 μV, 水平眼电HEOG的剔除阈限为± 80 μV。所有事件相关电位的分析只选取混合对中被试判断正确试次的EEG进行叠加和平均。
根据前人研究, 选取N2pc和SPCN为指标来反映注意定向和注意解除(Holmes et al., 2009; Holmes et al., 2014; Kappenman et al., 2014; Kappenman et al., 2015; Meconi et al., 2014)。选取头皮后部的P7/P8和PO7/PO8两对电极点的对侧与同侧电极波幅差值的均值计算N2pc和SPCN的幅值。由于用主观选择数据分析时间窗来计算平均波幅的方法可能存在较大的偏差, 因此我们选择成分的区域面积来度量成分(Sawaki, Geng, & Luck, 2012)。具体来说, N2pc波幅是180~300 ms内对侧减同侧的差异波中波幅为负值的区域的面积, SPCN则是300~600 ms内对侧减同侧的差异波中波幅为负值的区域的面积。由于采用面积来度量成分, 面积总是正数, 数据不符合正态分布, 不宜采用传统的参数检验, 我们采用更适合的非参数置换检验(non- permutation tests) (Gaspelin & Luck, 2018; Sawaki et al., 2012; Sun et al., 2018; Wang et al., 2016)。非参数置换检验的主要步骤是:首先随机编码威胁刺激试次的单侧性, 即将威胁刺激的同侧随机编码为对侧或同侧; 其次将随机编码试次后的EEG数据进行总平均, 并通过计算得到对侧减同侧的差异波; 最后从差异波中计算出对应时间窗内为负值的区域面积。以上步骤将进行1000次, 每次都计算出对应的值, 从而获得检验统计量的零分布(null distribution)。如果我们的实际观测波幅值大于检验统计量的零分布波幅值的95%, 则认为观测值反映了个体真实的生理反应而不是数据中的噪音。此外, 非参数置换检验还可用于检验组间数据是否存在差异, 其原理和主要步骤与条件之间的对比是一样的(Gaspelin & Luck, 2018; Sun et al., 2018; Wang et al., 2016)。在组间差异的非参数置换检验中, 对组别进行随机编码, 即将应激组随机编码为应激组或控制组, 随后计算组间差异值。以上步骤重复1000次, 每次都计算出对应的值, 从而获得组间差异数据的检验统计量的零分布。如果组间差异观测波幅值大于或小于检验统计量的零分布(双尾分布)波幅值的95%, 则认为组间差异具有显著的统计学意义。
3 结果
3.1 问卷
在特质焦虑分数上, 应激组(42.72 ± 4.40)和控制组(42.27 ± 5.54)没有显著差异, t(2, 34) = 0.27, p = 0.79。在状态焦虑分数上, 进行2(组别:应激组/控制组) × 5(测量时间:-85 min / -55 min / 0 min / 40 min / 70 min)重复测量方差分析, 结果显示(见图2(a))测量时间的主效应显著, F(4, 24) = 3.27, p = 0.046, η2p = 0.35; 组别主效应不显著, F(1, 27) = 0.83, p = 0.37; 组别和测量时间的交互作用不显著, F(4, 108) = 0.56, p = 0.60。由于具有比较明确的假设, 应激之后个体的状态焦虑会显著的提升, 我们对0 min (即社会评估冷压任务后)的状态焦虑进行了单独分析, 应激组(43.93 ± 1.12)显著高于控制组(40.21 ± 1.16), F(1, 27) = 5.33, p = 0.03, η2p = 0.16。同时, 在社会评估冷压任务前的-85 min (F(1, 27) = 0.83, p = 0.37)和-55 min (F(1, 27) = 0.39, p = 0.54)应激组和控制组在状态焦虑上均未发现显著差异, 结果说明SECPT任务显著提高了个体的状态焦虑。
图2
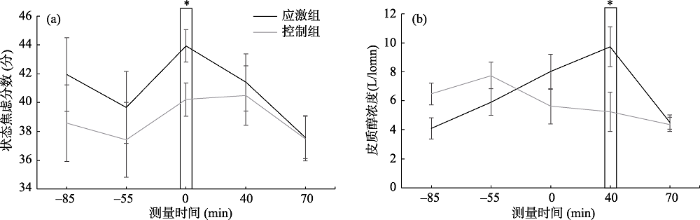
图2
应激组和控制组在不同时间点上的状态焦虑分数(a)和皮质醇浓度(b)结果。在社会评估冷压任务后, 应激组的状态焦虑(0 min)和皮质醇浓度(40 min)均显著高于控制组。
注:*表示p < 0.05。
3.2 皮质醇
在皮质醇浓度上, 进行2(组别:应激组/控制组) × 5(测量时间:-85 min / -55 min / 0 min / 40 min / 70 min)重复测量方差分析, 结果显示(见图2(b)), 测量时间的主效应显著, F(4, 31) = 5.71, p = 0.006, η2p = 0.42; 组别主效应不显著, F(1, 34) = 0.29, p = 0.60。组别和测量时间的交互作用显著, F(4, 136) = 7.50, p = 0.001, η2p = 0.18。进一步分析发现, 40 min应激组(9.74 ± 1.36)显著高于控制组(5.25 ± 1.36), F(1, 34) = 5.44, p = 0.03, η2p = 0.14; -85 min应激组(4.10 ± 0.75)显著低于控制组(6.47 ± 0.75), F(1, 34) = 5.01, p = 0.03, η2p = 0.13。
为了进一步确认应激任务诱发的有效性, 分别对应激组和控制组0 min和40 min时的皮质醇浓度进行t检验, 结果发现, 应激组在40 min (9.74 ± 7.81)的皮质醇浓度显著高于0 min (8.01 ± 6.96), t(1, 17) = -2.45, p = 0.03, Cohen’s d = 0.23; 控制组在40 min (5.25 ± 2.42)的皮质醇浓度与0 min (5.63 ± 2.19)无显著差异, t(1, 17) = 1.088, p = 0.292。结果说明SECPT导致应激组的皮质醇浓度显著提升。
3.3 行为学结果
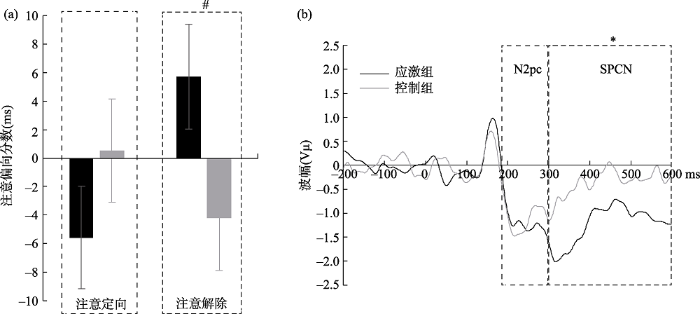
在行为学结果上, 对注意偏向分数的分析结果显示(见图3(a)), 组别主效应不显著, F(1, 34) = 0.85, p = 0.36; 注意偏向的主效应不显著, F(1, 34) = 0.49, p = 0.49; 组别与注意偏向的交互作用边缘显著, F(1, 34) = 2.87, p = 0.099。因此我们对两个组的注意定向和注意解除的分数进行单独的对比, 应激组与控制组在注意解除上存在边缘显著, 应激组(5.72 ± 3.69 ms)慢于控制组(-4.17 ± 3.69 ms), F(1, 34) = 3.59, p = 0.07, η2p = 0.10。在注意定向上两组无显著差异, F(1, 34) = 1.44, p = 0.24。
图3
3.4 ERP结果
3.4.1 N2pc
对N2pc的波幅进行独立样本t检验, 分析结果表明(见图3(b)), 应激组(0.14 ± 0.15)与控制组(0.13 ± 0.11)不存在显著差异, t(2, 34) = 0.17, p = 0.87。说明应激组和控制组在对威胁刺激上的注意定向没有差异。
图4

图4
应激组和控制组的置换检验结果(a)和组间差异(b)
注:(a)应激组和控制组中对侧减去同侧的差异波(相对于威胁刺激位置)的负值区域面积(N2pc和SPCN)。蓝色直方图表示1000次检验统计量的零分布。红色直线表示实际观测到的负值区域面积统计值。黄色区域表示检验统计量零分布的前5%。(b)负值区域面积统计值的组间差异(应激组减去控制组)。红色直线表示实际观测到的组间差异负值区域面积统计值。
3.4.2 SPCN
对SPCN的波幅进行独立样本t检验, 分析结果表明(见图3(b)), 应激组(0.42 ± 0.44)显著大于控制组(0.19 ± 0.13), t(2, 34) = 2.13, p = 0.04, Cohen’s d = 0.71。说明应激组和控制组在对威胁刺激的注意解除上具有显著差异。
3.5 相关性
为了进一步探讨急性应激对注意定向和注意解除的影响, 我们选取了应激组和控制组皮质醇增量的差异和N2pc、SPCN在两组上的差异分别进行相关分析。组间皮质醇增量差异为应激组的增量减去控制组的增量, 皮质醇增量为40 min和0 min时的皮质醇浓度差值。N2pc和SPCN的组间差异为应激组与控制组脑电成分时间窗内区域面积的差。结果发现, 组间皮质醇增量差异与N2pc的组间差异(r = 0.49, p = 0.04)和SPCN的组间差异(r = 0.47, p = 0.05)均具有显著的正相关关系。
3.6 时间效应
急性应激诱发的交感神经系统和HPA轴激活对脑认知功能的调节存在重要的时效性特征(de Kloet, Joels, & Holsboer, 2005; Hermans, Henckens, Joëls, & Fernández, 2014; Luo, Lin, Wu, & Qin, 2013)。因此,为了进一步探讨急性应激诱发的交感神经系统和HPA轴激活对注意定向和注意解除的不同影响, 我们将480个实验试次按照时间进程进行对半平分, 前半部分和后半部分实验试次各240个, 其中一致条件和不一致条件各90个, 中性对和混合对各30个。在行为学数据上, 进行2(组别:应激组/控制组) × 2(试次时间:前半部分/后半部分) × 2(注意偏向:注意定向/注意解除)的重复测量方差分析。结果发现, 组别主效应[F(1, 68) = 0.69, p = 0.41]、试次时间主效应[F(1, 68) = 0.01, p = 0.94]、注意偏向主效应[F(1, 34) = 0.46, p = 0.50]、组别、试次时间和注意偏向的三方交互作用[F(1, 34) = 0.37, p = 0.55]均不存在显著差异。
在ERP上, 分别对N2pc和SPCN进行2(组别:应激组/控制组) × 2(试次时间:前半部分/后半部分)的重复测量方差分析, 结果发现, 在N2pc上, 组别主效应[F(1, 34) = 0.005, p = 0.94]、试次时间主效应[F(1, 34) = 0.02, p = 0.90]均不存在显著差异, 组别和试次时间的交互作用显著, F(1, 34) = 9.23, p = 0.01, η2p = 0.21。但进一步分析发现, 应激组和控制组在前半部分[F(1, 34) = 2.63, p = 0.11]和后半部分[F(1, 34) = 2.06, p = 0.16]试次上的N2pc均不存在显著差异。在SPCN上, 组别主效应[F(1, 34) = 2.59, p = 0.12]、试次时间主效应[F(1, 34) = 0.01, p = 0.91]均不存在显著差异, 组别和试次时间的交互作用显著, F(1, 34) = 3.94, p = 0.06, η2p = 0.10。进一步分析发现, 应激组和控制组在前半部分试次上的SPCN无显著差异, F(1, 34) = 0.02, p = 0.90; 在后半部分试次上的SPCN应激组(0.27 ± 0.06)显著高于控制组(0.09 ± 0.06), F(1, 34) = 4.92, p = 0.03, η2p = 0.13。结果表明, 急性应激对注意偏向的影响主要在点探测任务中实验试次的后半部分, 这与HPA轴的激活时间比较接近, 说明急性应激对注意偏向的影响主要是由HPA轴激活导致的, 即HPA轴激活会损害个体对威胁刺激的注意解除, 这与本研究中前面的发现是一致的。
4 讨论
本研究主要探讨急性应激对威胁刺激注意定向和注意解除的影响。研究发现, SECPT任务显著的诱发了个体的应激体验。SECPT任务结束后应激组的状态焦虑分数和皮质醇浓度显著高于控制组。在注意偏向中, 应激组和控制组在注意定向上无显著差异, 在注意解除上应激组慢于控制组。ERP结果上, 应激组和控制组在N2pc上无显差异; 在SPCN上, 应激组比控制组产生更负的波幅。在急性应激与注意定向和注意解除的相关性结果上, 组间皮质醇增量差异值和N2pc、SPCN的组间差异值均具有显著的正相关。在时间效应上, 在后半部分试次上应激组比控制组的SPCN更负。结果表明急性应激会影响个体对威胁刺激的注意偏向, 主要表现为急性应激损害了个体对威胁刺激的注意解除, 使个体对威胁刺激的解除变得更困难, 在威胁刺激上的注意维持时间更长。
皮质醇浓度和主观状态评价的研究结果表明, SECPT任务有效地诱发了个体的应激体验。在皮质醇浓度上, 应激组的皮质醇浓度显著高于控制组。先前研究表明, SECPT任务后25分钟左右个体的唾液皮质醇浓度达到峰值, 且应激效果能持续 35~60分钟左右(Luo et al., 2018; Schwabe et al., 2008; Schwabe, Höffken, Tegenthoff, & Wolf, 2013; Schwabe, Joëls, Roozendaal, Wolf, & Oitzl, 2012; Schwabe & Schächinger, 2018), 这与本研究中SECPT任务结束后40分钟的皮质醇浓度的测量结果基本一致。但本研究中在SECPT任务开始前85分钟(-85 min)时应激组的皮质醇浓度显著低于控制组, 其原因可能是由于个体对实验室环境适应性差异所引起。我们对个体到达实验室休息放松20分钟后(-55 min)时的皮质醇浓度进行进一步分析, 结果发现应激组与控制组无显著差异, 说明个体适应实验室环境后皮质醇浓度回到正常水平, 因此-85 min时应激组和控制组的皮质醇浓度结果并不会影响应激诱发的结果。从主观状态评价来看, 应激组的焦虑水平显著高于控制组。Garrett等人使用状态焦虑问卷评估应激环境中个体的情绪体验, 结果发现个体的状态焦虑得分与个体的应激体验具有显著的相关(Garrett, González-Garzón, Foulkes, Levita, & Sharot, 2018), 与本研究中SECPT任务结束后(0 min)应激组的状态焦虑得分显著高于控制组的结果基本一致。与以往研究不一致的是我们没有观察到组别与测量时间的交互作用, 可能是由于在SECPT任务之前的时间点上, 应激组的状态焦虑分数高于控制组, 但在数理统计上并没有发现显著的差异。
注意偏向和ERP的研究结果发现, 急性应激并不会增强个体对威胁刺激的注意定向, 而是使个体对威胁刺激的注意解除变得更困难, 在威胁刺激上的注意维持时间更长。在注意偏向中, 应激组和控制组在注意定向上无显著差异, 在注意解除上应激组慢于控制组。本研究中控制组的研究结果与Armstrong等人的研究一致。Armstrong等人研究发现, 在视觉搜索任务中个体对威胁刺激会维持更长的注意时间更长, 从而导致个体对威胁刺激的注意解除更慢(Armstrong & Olatunji, 2012)。本研究中, SECPT任务有效地诱发了个体的焦虑状态, 从而损害了个体对威胁刺激的注意解除能力。对与急性应激相关疾病(如:焦虑症、抑郁症)的研究也发现, 高焦虑的个体更难将注意力从威胁刺激中解除出来(Taylor, Cross, & Amir, 2016)。同时, 抑郁个体不会更快地对威胁刺激(如:悲伤的面孔)进行注意定向(Cisler & Koster, 2010), 但抑郁个体会在威胁刺激出现较短时间后将注意力集中在威胁刺激上(Peckham, McHugh, & Otto, 2010)。此外, 先前研究发现, 威胁刺激的呈现时间也会影响个体对威胁刺激的注意偏向。当威胁刺激的呈现时间很短(如:≤100 ms)时, 个体往往会对威胁刺激表现出注意定向加速; 而当威胁刺激呈现时间相对较长(如:≥500 ms)时, 往往表现为注意解除困难(Cooper & Langton, 2006)。本研究中威胁刺激在400~600 ms内随机呈现时, 个体对威胁刺激表现出注意解除困难, 但在注意定向上没有差异, 与先前研究基本一致。
在ERP结果上, 应激组和控制组在N2pc上无显著差异, 应激组的SPCN比控制组更负。同时, 组间皮质醇增量差异和N2pc、SPCN的组间差异均具有显著的正相关。先前研究发现, N2pc反映了个体对刺激的注意转移, SPCN反映的是个体对信息的注意维持(Holmes et al., 2009; Holmes et al., 2014; Meconi et al., 2014)。本研究中, 应激组和控制组在N2pc上没有显著差异, 我们推测其原因可能是:在信息加工的早期阶段, 威胁刺激呈现的时间较长, 急性应激抑制了个体早期对威胁刺激的注意定向。先前研究发现, 杏仁核是加工恐惧相关信息以及表达恐惧相关行为的关键大脑结构(LeDoux, 2003)。在急性应激状态下, 个体分泌过多的皮质醇会改变杏仁核和额顶网络之间的功能连接, 削弱杏仁核对威胁刺激的特异性响应, 改变个体对威胁刺激的神经机制, 使个体在早期对威胁刺激的加工由特异性转向敏感性, 对刺激不加区分或辨别的选择性反应, 威胁刺激和中性刺激将具有类似的认知加工过程, 并且具有类似的ERP成分响应(Grant, Judah, White, & Mills, 2015; Sänger et al., 2014; van Marle, Hermans, Qin, & Fernández, 2009), 这与组间皮质醇增量差异和N2pc的组间差异具有显著正相关的结果基本一致。因此, 急性应激影响了个体对威胁刺激的反应但并没有显著增加对威胁刺激的注意定向。但应激组的SPCN比控制组更负,表明在确定了威胁刺激之后应激组比控制组将更多的注意资源维持在威胁刺激上。Weymar等人研究发现急性应激下威胁刺激(如:不愉悦的图片)会诱发更大且持续时间更久的LPP成分, 说明急性应激会强化个体对威胁性的注意维持(Weymar et al., 2012)。Meconi等人对焦虑个体的研究也发现, 与低焦虑个体相比, 威胁刺激(如:不值得信任的面孔)会诱发高焦虑个体更负的SPCN, 说明高焦虑个体会对威胁刺激进行持续地注意加工(Meconi et al., 2014), 这与本研究中急性应激诱发的HPA轴激活会损害个体对威胁刺激注意解除的结果一致。Taylor等人研究发现, 注意控制能力会调节焦虑个体对威胁刺激的注意解除能力, 注意控制能力越弱的个体对威胁刺激的注意解除速度越慢, 反之则相反(Taylor et al., 2016)。Gindt等人的研究也发现了类似的结果, Gindt等人研究发现创伤后应激障碍个体比焦虑症个体更难从威胁刺激中解除出来(Gindt, Nachon, Chanquoy, Faure, & Garcia, 2017)。先前研究发现, 额-顶网络主要负责注意资源的分配(Fecteau & Munoz, 2006; Gottlieb, 2007)。急性应激损害个体对威胁刺激的注意解除的神经机制, 可能是急性应激损害了额-顶网络与注意解除相关的脑区功能, 导致背外侧前额叶和其他脑区的功能连接性降低,个体未能有效的控制注意力分配, 从而使个体自上而下的注意控制资源重新分配会减少或消失(Luo et al., 2018; Qin et al., 2009; Sänger et al., 2014; Weerda, Muehlhan, Wolf, & Thiel, 2010; Wu et al., 2014), 这与组间皮质醇增量差异与SPCN的组间差异具有显著正相关的结果一致。因此, 急性应激会损害个体对威胁刺激的注意解除, 在对威胁刺激加工晚期产生更负的SPCN。
5 结论
急性应激对个体在威胁刺激的注意定向上没有显著影响, 但会损害个体对威胁刺激的注意解除。其原因可能是因为急性应激损害了与注意解除相关的额-顶网络的功能。
参考文献
Eye tracking of attention in the affective disorders: A meta-analytic review and synthesis
DOI:10.1016/j.cpr.2012.09.004
URL
[本文引用: 1]

A large body of research has demonstrated that affective disorders are characterized by attentional biases for emotional stimuli. However, this research relies heavily on manual reaction time (RT) measures that cannot fully delineate the time course and components of attentional bias. Eye tracking technology, which allows relatively direct and continuous measurement of overt visual attention, may provide an important supplement to RT measures. This article reviews eye tracking research on anxiety and depression, evaluating the experimental paradigms and eye movement indicators used to study attentional biases. Also included is a meta-analysis of extant eye tracking research (33 experiments; N=1579) on both anxiety and depression. Relative to controls, anxious individuals showed increased vigilance for threat during free viewing and visual search, and showed difficulty disengaging from threat in visual search tasks, but not during free viewing. In contrast, depressed individuals were not characterized by vigilance for threat during free viewing, but were characterized by reduced orienting to positive stimuli, as well as reduced maintenance of gaze on positive stimuli and increased maintenance of gaze on dysphoric stimuli. Implications of these findings for theoretical accounts of attentional bias in anxiety and depression are discussed, and avenues for future research using eye-tracking technology are outlined. (C) 2012 Elsevier Ltd.
Stress signalling pathways that impair prefrontal cortex structure and function
DOI:10.1038/nrn2648
URL
PMID:19455173
[本文引用: 2]
The prefrontal cortex (PFC) - the most evolved brain region - subserves our highest-order cognitive abilities. However, it is also the brain region that is most sensitive to the detrimental effects of stress exposure. Even quite mild acute uncontrollable stress can cause a rapid and dramatic loss of prefrontal cognitive abilities, and more prolonged stress exposure causes architectural changes in prefrontal dendrites. Recent research has begun to reveal the intracellular signalling pathways that mediate the effects of stress on the PFC. This research has provided clues as to why genetic or environmental insults that disinhibit stress signalling pathways can lead to symptoms of profound prefrontal cortical dysfunction in mental illness.
How many trials does it take to get a significant ERP effect? It depends
DOI:10.1111/psyp.13049
URL
PMID:29266241
[本文引用: 1]
In designing an ERP study, researchers must choose how many trials to include, balancing the desire to maximize statistical power and the need to minimize the length of the recording session. Recent studies have attempted to quantify the minimum number of trials needed to obtain reliable measures for a variety of ERP components. However, these studies have largely ignored other variables that affect statistical power in ERP studies, including sample size and effect magnitude. The goal of the present study was to determine whether and how the number of trials, number of participants, and effect magnitude interact to influence statistical power, thus providing a better guide for selecting an appropriate number of trials. We used a Monte Carlo approach to measure the probability of obtaining a statistically significant result when testing for (a) the presence of an ERP effect, (b) within-participant condition differences in an ERP effect, and (c) between-participants group differences in an ERP effect. Each of these issues was examined in the context of the error-related negativity and the lateralized readiness potential. We found that doubling the number of trials recommended by previous studies led to more than a doubling of statistical power under many conditions. Thus, when determining the number of trials that should be included in a given study, researchers must consider the sample size, the anticipated effect magnitude, and the noise level, rather than relying solely on general recommendations about the number of trials needed to obtain a &quot;stable&quot; ERP waveform.
Nonconscious attention bias to threat is correlated with anterior cingulate cortex gray matter volume: A voxel- based morphometry result and replication
DOI:10.1016/j.neuroimage.2011.09.040
URL
[本文引用: 2]
An important aspect of the fear response is the allocation of spatial attention toward threatening stimuli. This response is so powerful that modulations in spatial attention can occur automatically without conscious awareness. Functional neuroimaging research suggests that the amygdala and anterior cingulate cortex (ACC) form a network involved in the rapid orienting of attention to threat. A hyper-responsive attention bias to threat is a common component of anxiety disorders. Yet, little is known of how individual differences in underlying brain morphometry relate to variability in attention bias to threat. Here, we performed two experiments using dot-probe tasks that measured individuals' attention bias to backward masked fearful faces. We collected whole-brain structural magnetic resonance images and used voxel-based morphometry to measure brain morphometry. We tested the hypothesis that reduced gray matter within the amygdala and ACC would be associated with reduced attention bias to threat. In Experiment 1, we found that backward masked fearful faces captured spatial attention and that elevated attention bias to masked threat was associated with greater ACC gray matter volumes. In Experiment 2, this association was replicated in a separate sample. Thus, we provide initial and replicating evidence that ACC gray matter volume is correlated with biased attention to threat. Importantly, we demonstrate that variability in affective attention bias within the healthy population is associated with ACC morphometry. This result opens the door for future research into the underlying brain morphometry associated with attention bias in clinically anxious populations. (C) 2011 Elsevier Inc.
Masked fearful faces modulate the orienting of covert spatial attention
DOI:10.1037/a0012653
URL
PMID:18729582
[本文引用: 3]
Dot probe studies indicate that masked fearful faces modulate spatial attention. However, without a baseline to compare congruent and incongruent reaction times, it is unclear which aspect(s) of attention (orienting or disengagement) is affected. Additionally, backward masking studies commonly use a neutral face as the mask stimulus. This method results in greater perceptual inconsistencies for fearful as opposed to neutral faces. Therefore, it is currently unclear whether the effects of backward masked fearful faces are due to the fearful nature of the face or perceptual inconsistencies. Equally unclear, is whether this spatial attention effect is due to orienting or disengagement. Two modified dot probe experiments with neutral (closed mouth in Experiment 1) and smiling (open mouth in Experiment 2) masks were used to determine the role of perceptual inconsistencies in mediating the spatial attention effects elicited by masked fearful faces. The results indicate that masked fearful faces modulate the orienting of spatial attention, and it appears that this effect is due to the fearful nature of the face rather than perceptual inconsistencies between the initial faces and masks.
Mechanisms of attentional biases towards threat in anxiety disorders: An integrative review
DOI:10.1016/j.cpr.2009.11.003
URL
[本文引用: 1]
AbstractA wealth of research demonstrates attentional biases toward threat in the anxiety disorders. Several models have been advanced to explain these biases in anxiety, yet the mechanisms comprising and mediating the biases remain unclear. In the present article, we review evidence regarding the mechanisms of attentional biases through careful examination of the components of attentional bias, the mechanisms underlying these components, and the stage of information processing during which the biases occur. Facilitated attention, difficulty in disengagement, and attentional avoidance comprise the components of attentional bias. A threat detection mechanism likely underlies facilitated attention, a process that may be neurally centered around the amygdala. Attentional control ability likely underlies difficulty in disengagement, emotion regulation goals likely underlie attentional avoidance, and both of these processes may be neurally centered around prefrontal cortex functioning. The threat detection mechanism may be a mostly automatic process, attentional avoidance may be a mostly strategic process, and difficulty in disengagement may be a mixture of automatic and strategic processing. Recommendations for future research are discussed.]]>
Attentional bias to angry faces using the dot-probe task? It depends when you look for it
DOI:10.1016/j.brat.2005.10.004
URL
[本文引用: 1]
AbstractA number of studies using the dot-probe task now report the existence of an attentional bias to angry faces in participants who rate highly on scales of anxiety; however, no equivalent bias has been observed in non-anxious populations, despite evidence to the contrary from studies using other tasks. One reason for this discrepancy may be that researchers using the dot-probe task have rarely investigated any effects which might emerge earlier than 500 ms following presentation of the threat-related faces. Accordingly, in the current study we presented pairs of face stimuli with emotional and neutral expressions and probed the allocation of attention to these stimuli for presentation times of 100 and 500 ms. Results showed that at 100 ms there was an attentional bias towards the location of the relatively threatening stimulus (the angry face in angry/neutral pairs and the neutral face in neutral/happy pairs) and this pattern reversed by 500 ms. Comparisons of reaction time (RT) scores with an appropriate baseline suggested that the early bias toward threatening faces may actually arise through inhibition of the relatively least threatening member of a face pair rather than through facilitation of, or vigilance towards, the more threatening stimulus. However the mechanisms governing the observed biases are interpreted, these data provide evidence that probing for the location of spatial attention at 500 ms is not necessarily indicative of the initial allocation of attention between competing emotional facial stimuli.]]>
Acute stress modulates genotype effects on amygdala processing in humans
DOI:10.1073/pnas.1003514107
URL
PMID:20457919
[本文引用: 2]
Probing gene-environment interactions that affect neural processing is crucial for understanding individual differences in behavior and disease vulnerability. Here, we tested whether the current environmental context, which affects the acute brain state, modulates genotype effects on brain function in humans. We manipulated the context by inducing acute psychological stress, which increases noradrenergic activity, and probed its effect on tonic activity and phasic responses in the amygdala using two MRI techniques: conventional blood oxygen level-dependent functional MRI and arterial spin labeling. We showed that only carriers of a common functional deletion in ADRA2B, the gene coding for the alpha2b-adrenoreceptor, displayed increased phasic amygdala responses under stress. Tonic activity, reflecting the perfusion of the amygdala, increased independently of genotype after stress induction. Thus, when tonic activity was heightened by stress, only deletion carriers showed increased amygdala responses. Our results demonstrate that genetic effects on brain operations can be state dependent, such that they only become apparent under specific, often environmentally controlled, conditions.
Noradrenergic signaling in the amygdala contributes to the reconsolidation of fear memory: Treatment implications for PTSD
DOI:10.1196/annals.1364.056
URL
PMID:16891611
[本文引用: 1]
Intrusive memories resulting from an emotional trauma are a defining feature of posttraumatic stress disorder (PTSD). Existing studies demonstrate that an increase of noradrenergic activity during a life-threatening event contributes to strengthening or &quot;overconsolidation&quot; of the memory for trauma. The lateral nucleus of the amygdala (LA) is critical for fear learning. Using classical fear conditioning in rats, we have recently demonstrated that noradrenergic blockade in the LA following reactivation of fear memory by retrieval disrupts memory reconsolidation and lastingly impairs fear memory. This suggests that noradrenergic blockade may be useful in attenuating traumatic memories, even well-consolidated old memories, in PTSD.
Stress and the brain: From adaptation to disease
DOI:10.1038/nrn1683
URL
PMID:15891777
[本文引用: 1]
In response to stress, the brain activates several neuropeptide-secreting systems. This eventually leads to the release of adrenal corticosteroid hormones, which subsequently feed back on the brain and bind to two types of nuclear receptor that act as transcriptional regulators. By targeting many genes, corticosteroids function in a binary fashion, and serve as a master switch in the control of neuronal and network responses that underlie behavioural adaptation. In genetically predisposed individuals, an imbalance in this binary control mechanism can introduce a bias towards stress-related brain disease after adverse experiences. New candidate susceptibility genes that serve as markers for the prediction of vulnerable phenotypes are now being identified.
Salience, relevance, and firing: A priority map for target selection
DOI:10.1016/j.tics.2006.06.011
URL
PMID:16843702
[本文引用: 3]
The salience map is a crucial concept for many theories of visual attention. On this map, each object in the scene competes for selection - the more conspicuous the object, the greater its representation, and the more likely it will be chosen. In recent years, the firing patterns of single neurons have been interpreted using this framework. Here, we review evidence showing that the expression of salience is remarkably similar across structures, remarkably different across tasks, and modified in important ways when the salient object is consistent with the goals of the participant. These observations have important ramifications for theories of attention. We conclude that priority--the combined representation of salience and relevance--best describes the firing properties of neurons.
Menstrual cycle-dependent neural plasticity in the adult human brain is hormone, task, and region specific
In rodents, cyclically fluctuating levels of gonadal steroid hormones modulate neural plasticity by altering synaptic transmission and synaptogenesis. Alterations of mood and cognition observed during the menstrual cycle suggest that steroid-related plasticity also occurs in humans. Cycle phase-dependent differences in cognitive performance have almost exclusively been found in tasks probing lateralized neuronal domains, i.e., cognitive domains such as language, which are predominantly executed by one hemisphere. To search for neural correlates of hormonally mediated neural plasticity in humans, we thus conducted a functional magnetic resonance imaging study measuring brain activity related to a semantic decision task in the language domain. This was contrasted with a letter-matching task in the perceptual domain, in which we expected no steroid hormone-mediated effect. We investigated 12 young healthy women in a counterbalanced repeated-measure design during low-steroid menstruation and high-steroid midluteal phase. Steroid serum levels correlated with the volume and lateralization of particular brain activations related to the semantic task but not with brain activity related to the perceptual task. More specifically, bilateral superior temporal recruitment correlated positively with progesterone and medial superior frontal recruitment with both progesterone and estradiol serum levels, whereas activations in inferior and middle frontal cortex were unaffected by steroid levels. In contrast to these specific interactions, testosterone levels correlated nonselectively with overall activation levels by neural and/or vascular factor(s). In conclusion, our data demonstrate steroid hormone responsivity in the adult human brain by revealing neural plasticity in the language domain, which appears hormone, task, and region specific.
Do threatening stimuli draw or hold visual attention in subclinical anxiety?
Biases in information processing undoubtedly play an important role in the maintenance of emotion and emotional disorders. In an attentional cueing paradigm, threat words and angry faces had no advantage over positive or neutral words (or faces) in attracting attention to their own location, even for people who were highly state-anxious. In contrast, the presence of threatening cues (words and faces) had a strong impact on the disengagement of attention. When a threat cue was presented and a target subsequently presented in another location, high state-anxious individuals took longer to detect the target relative to when either a positive or a neutral cue was presented. It is concluded that threat-related stimuli affect attentional dwell time and the disengage component of attention, leaving the question of whether threat stimuli affect the shift component of attention open to debate.
Updating beliefs under perceived threat
DOI:10.1523/JNEUROSCI.0716-18.2018
URL
PMID:30082420
[本文引用: 1]
SIGNIFICANCE STATEMENT The human tendency to be overly optimistic has mystified scholars and lay people for decades: How could biased beliefs have been selected over unbiased beliefs? Scholars have suggested that although the optimism bias can lead to negative outcomes, including financial collapse and war, it can also facilitate health and productivity. Here, we demonstrate that a mechanism generating the optimism bias, namely asymmetric information integration, evaporates under threat. Such flexibility could result in enhanced caution in dangerous environments while supporting an optimism bias otherwise, potentially increasing well-being.]]>
Combined electrophysiological and behavioral evidence for the suppression of salient distractors
DOI:10.1162/jocn_a_01279
URL
PMID:29762104
[本文引用: 2]
D) component, a putative neural index of suppression. Other support comes from a handful of behavioral studies showing that processing at the salient locations is inhibited compared with other locations. The current study was designed to link the behavioral and neural evidence by combining ERP recordings with an experimental paradigm that provides a behavioral measure of suppression. We found that, when a salient distractor item elicited the PD component, processing at the location of this distractor was suppressed below baseline levels. Furthermore, the magnitude of behavioral suppression and the magnitude of the PD component covaried across participants. These findings provide a crucial connection between the behavioral and neural measures of suppression, which opens the door to using the PD component to assess the timing and neural substrates of the behaviorally observed suppression.]]>
Attentional bias in post-traumatic stress symptoms or anxiety
DOI:10.1007/s00068-017-0808-4
URL
PMID:28634651
[本文引用: 1]
Recent studies have demonstrated that black patients receive substandard care compared with white patients across healthcare settings. The purpose of this study was to evaluate the association of race on the management (salvage vs. amputation) of traumatic lower extremity open fractures.
From thought to action: The parietal cortex as a bridge between perception, action, and cognition
DOI:10.1016/j.neuron.2006.12.009
URL
[本文引用: 3]
binds visuospatial, motor, and cognitive information into a topographically organized signal of behavioral salience. By specifying attentional priority as a synthesis of multiple task demands, LIP operates at the interface of perception, action, and cognition.]]>
Worry and discrimination of threat and safety cues: An event-related potential investigation
DOI:10.1016/j.beth.2014.09.015
URL
PMID:26459845
[本文引用: 1]
Overgeneralization of fear to safety cues is increasingly being studied in order to further our understanding of the maintenance of anxiety disorders. The current study used event-related potentials (ERPs) to evaluate whether worry affects anticipation and processing of threat and neutral pictures during a conditioning task. Fifty-two high (n=24) and low (n=28) worriers completed a paradigm in which a neutral stimulus indicated the valence of a second stimulus, either a threat or neutral picture. Results found that worriers displayed reduced anticipatory responses to both stimulus types as indexed by the stimulus preceding negativity, although they displayed an increased stimulus preceding negativity to threatening images during the second half of the task. In addition, high and low worriers differed in processing of threat and neutral images as indexed by the late positive potential. These findings support the overgeneralization of fear literature, suggesting that worriers display difficulty discriminating safety cues from threat cues, and this affects the attentional resources devoted to subsequent stimuli. Implications of these results are discussed.
Dynamic adaptation of large-scale brain networks in response to acute stressors
DOI:10.1016/j.tins.2014.03.006
URL
[本文引用: 1]
Stress initiates an intricate response that affects diverse cognitive and affective domains, with the goal of improving survival chances in the light of changing environmental challenges. Here, we bridge animal data at cellular and systems levels with human work on brain-wide networks to propose a framework describing how stress-related neuromodulators trigger dynamic shifts in network balance, enabling an organism to comprehensively reallocate its neural resources according to cognitive demands. We argue that exposure to acute stress prompts a reallocation of resources to a salience network, promoting fear and vigilance, at the cost of an executive control network. After stress subsides, resource allocation to these two networks reverses, which normalizes emotional reactivity and enhances higher-order cognitive processes important for long-term survival.
Stress-related noradrenergic activity prompts large-scale neural network reconfiguration
DOI:10.1126/science.1209603
URL
[本文引用: 1]
Acute stress shifts the brain into a state that fosters rapid defense mechanisms. Stress-related neuromodulators are thought to trigger this change by altering properties of large-scale neural populations throughout the brain. We investigated this brain-state shift in humans. During exposure to a fear-related acute stressor, responsiveness and interconnectivity within a network including cortical (frontoinsular, dorsal anterior cingulate, inferotemporal, and temporoparietal) and subcortical (amygdala, thalamus, hypothalamus, and midbrain) regions increased as a function of stress response magnitudes. beta-adrenergic receptor blockade, but not cortisol synthesis inhibition, diminished this increase. Thus, our findings reveal that noradrenergic activation during acute stress results in prolonged coupling within a distributed network that integrates information exchange between regions involved in autonomic-neuroendocrine control and vigilant attentional reorienting.
Attentional selectivity for emotional faces: Evidence from human electrophysiology
DOI:10.1111/j.1469-8986.2008.00750.x
URL
PMID:19055500
[本文引用: 7]
This study investigated the temporal course of attentional biases for threat-related (angry) and positive (happy) facial expressions. Electrophysiological (event-related potential) and behavioral (reaction time [RT]) data were recorded while participants viewed pairs of faces (e.g., angry face paired with neutral face) shown for 500 ms and followed by a probe. Behavioral results indicated that RTs were faster to probes replacing emotional versus neutral faces, consistent with an attentional bias for emotional information. Electrophysiological results revealed that attentional orienting to threatening faces emerged earlier (early N2pc time window; 180-250 ms) than orienting to positive faces (after 250 ms), and that attention was sustained toward emotional faces during the 250-500-ms time window (late N2pc and SPCN components). These findings are consistent with models of attention and emotion that posit rapid attentional prioritization of threat.
Electrophysiological evidence for greater attention to threat when cognitive control resources are depleted
DOI:10.3758/s13415-013-0212-4
URL
PMID:24165903
[本文引用: 7]
In this study, we investigated the time course of attentional bias for threat-related (angry) facial expressions under conditions of high versus low cognitive (working memory) load. Event-related potential (ERP) and reaction time (RT) data were recorded while participants viewed pairs of faces (angry paired with neutral face) displayed for 500 ms and followed by a probe. Participants were required to respond to the probe while performing a concurrent task of holding in working memory a sequence of digits that were either in the same order (low memory load) or in a random mixed order (high memory load). The ERP results revealed that higher working memory load resulted in enhanced lateralized neural responses to threatening relative to neutral faces, consistent with greater initial orienting of attention to threatening faces (early N2pc: 180-252 ms) and enhanced maintenance of processing representations of threat (late N2pc, 252-320 ms; SPCN, 320-500 ms). The ERP indices showed significant positive relationships with each other, and also with the behavioral index of attentional bias to threat (reflected by faster RTs to probes replacing angry than neutral faces at 500 ms), although the latter index was not significantly influenced by memory load. Overall, the findings indicate that depletion of cognitive control resources, using a working memory manipulation, increases the capacity of task-irrelevant threat cues to capture and hold attention.
Acute psychological stress disrupts attentional bias to threat-related stimuli
DOI:10.1038/s41598-017-14138-w
URL
PMID:29097660
[本文引用: 1]
The present study investigated the effect of acute stress on attentional bias to threat using behavioral and ERP methods. Sixty-two male participants were randomly assigned to a stress condition (Trier Social Stress Test) or a control condition. To examine the impact of stress-induced cortisol on attentional bias to threat, participants in the stress group were split into Low- and High cortisol responders. All participants were then administered a modified dot probe task in which the cues were neutral and angry faces. Behavioral results showed a pattern of attentional bias toward threat in the Control group but not in the stress group. For the ERPs, the P100 peaked earlier for the angry-cued targets than the neutral-cued targets in the Control group, which suggests a rapid, adaptive response toward threat. However, this effect was not observed in the stress group, suggesting a suppressed attentional bias under stress. In addition, the stress group (including both Low and High cortisol responders) showed reduced P300 amplitude to target onset than the Control group. These results suggest that acute stress disrupts attentional bias to threat including a reduction in early bias to threat in addition to a subsequent change of attention allocation.
Behavioral and ERP measures of attentional bias to threat in the dot-probe task: Poor reliability and lack of correlation with anxiety
DOI:10.3389/fpsyg.2014.01368
URL
PMID:25538644
[本文引用: 4]
The dot-probe task is often considered a gold standard in the field for investigating attentional bias to threat. However, serious issues with the task have been raised. Specifically, a number of studies have demonstrated that the traditional reaction time (RT) measure of attentional bias to threat in the dot-probe task has poor internal reliability and poor test-retest reliability. In addition, although threatening stimuli capture attention in other paradigms, attentional bias to threat has not usually been found in typical research participants in the dot-probe task. However, when attention is measured in the dot-probe task with the N2pc component of the event-related potential waveform, substantial attentional orienting to threat is observed, and the internal reliability is moderate. To provide a rigorous comparison of the reliability of this N2pc measure and the conventional behavioral measure, as well as to examine the relationship of these measures to anxiety, the present study examined the N2pc in conjunction with RT in the dot-probe task in a large sample of participants (N = 96). As in previous studies, RT showed no bias to threatening images across the sample and exhibited poor internal reliability. Moreover, this measure did not relate to trait anxiety. By contrast, the N2pc revealed a significant initial shift of attention to threat, and this measure was internally reliable. However, the N2pc was not correlated with trait anxiety, indicating that it does not provide a meaningful index of individual differences in anxiety in the dot-probe task. Together, these results indicate a serious need to develop new tasks and methods to more reliably investigate attentional bias to threat and its relationship to anxiety in both clinical and non-clinical populations.
Electrocortical evidence for rapid allocation of attention to threat in the dot-probe task
DOI:10.1093/scan/nsu098
URL
PMID:25062842
[本文引用: 4]
Threatening stimuli have been shown to preferentially capture attention using a range of tasks and measures. However, attentional bias to threat has not typically been found in unselected individuals using behavioral measures in the dot-probe task, one of the most common ways of examining attention to threat. The present study leveraged event-related potentials (ERPs) in conjunction with behavioral measures in the dot-probe task to examine whether more direct measures of attention might reveal an attentional bias to threat in unselected individuals. As in previous dot-probe studies, we found no evidence of an attentional bias to threat using reaction time; additionally, this measure exhibited poor internal reliability. In contrast, ERPs revealed an initial shift of attention to threat-related stimuli, reflected by the N2pc, which showed moderate internal reliability. However, there was no evidence of sustained engagement with the threat-related stimuli, as measured by the late positive potential (LPP). Together, these results demonstrate that unselected individuals do initially allocate attention to threat in the dot-probe task, and further, that this bias is better characterized by neural measures of attention than traditional behavioral measures. These results have implications for the study of attention to threat in both unselected and anxious populations.
Stress revisited: A critical evaluation of the stress concept
DOI:10.1016/j.neubiorev.2011.02.003
URL
[本文引用: 1]
AbstractWith the steadily increasing number of publications in the field of stress research it has become evident that the conventional usage of the stress concept bears considerable problems. The use of the term ‘stress’ to conditions ranging from even the mildest challenging stimulation to severely aversive conditions, is in our view inappropriate. Review of the literature reveals that the physiological ‘stress’ response to appetitive, rewarding stimuli that are often not considered to be stressors can be as large as the response to negative stimuli. Analysis of the physiological response during exercise supports the view that the magnitude of the neuroendocrine response reflects the metabolic and physiological demands required for behavioural activity. We propose that the term ‘stress’ should be restricted to conditions where an environmental demand exceeds the natural regulatory capacity of an organism, in particular situations that include unpredictability and uncontrollability. Physiologically, stress seems to be characterized by either the absence of an anticipatory response (unpredictable) or a reduced recovery (uncontrollable) of the neuroendocrine reaction. The consequences of this restricted definition for stress research and the interpretation of results in terms of the adaptive and/or maladaptive nature of the response are discussed.Research highlights? The review evaluates the stress response during rewarding and aversive conditions. ? A stress response is characterized by a reduced recovery rather than its magnitude. ? The concepts of adaptive capacity and regulatory range are introduced. ? Implications for the use of the stress concept in stress research are discussed.]]>
Selective attention to threat in the dot probe paradigm: Differentiating vigilance and difficulty to disengage
DOI:10.1016/j.brat.2003.08.001
URL
[本文引用: 1]
AbstractThe dot probe task [J Abnorm Psychol 95 (1986) 15] is an often-used paradigm to investigate selective attention to threat. A facilitated response to probes that appear at the same location of threat information in comparison with responses to probes at the opposite location of threat information is interpreted as vigilance for threat. We argue that the findings in the dot probe paradigm are ambiguous evidence for the vigilance to threat hypothesis. Results can also be interpreted as a difficulty to disengage from threat. In this study, 44 undergraduates performed a pictorial version of the probe detection task. Taking into account the reaction times on neutral trials, we found no evidence for a facilitated detection of threatening information. It was found that the dot probe effects are at least partially due to disengagement effects. The implications of these results for the understanding of attentional bias in normal and anxious individuals are discussed.]]>
Components of attentional bias to threat in high trait anxiety: Facilitated engagement, impaired disengagement, and attentional avoidance
DOI:10.1016/j.brat.2005.12.011
URL
[本文引用: 1]
AbstractThere is a wealth of evidence demonstrating enhanced attention to threat in high trait anxious individuals (HTA) compared with low trait anxious individuals (LTA). In two experiments, we investigated whether this attentional bias is related to facilitated attentional engagement to threat or difficulties disengaging attention from threat. HTA and LTA undergraduates performed a modified exogenous cueing task, in which the location of a target was correctly or incorrectly cued by neutral, highly and mildly threatening pictures. Results indicate that at 100 ms picture presentation, HTA individuals more strongly engaged their attention with and showed impaired disengagement from highly threatening pictures than LTA individuals. In addition, HTA individuals showed a stronger tendency to attentional avoidance of threat at the 200 and 500 ms picture presentation. These data provide evidence for differential patterns of anxiety-related biases in attentive processing of threat at early versus later stages of information processing.]]>
The emotional brain, fear, and the amygdala
DOI:10.1023/A:1025048802629
URL
[本文引用: 1]
When threat is near, get out of here: Dynamics of defensive behavior during freezing and active avoidance
DOI:10.1177/0956797615597332
URL
PMID:26408036
[本文引用: 1]
When detecting a threat, humans and other animals engage in defensive behaviors and supporting physiological adjustments that vary with threat imminence and potential response options. In the present study, we shed light on the dynamics of defensive behaviors and associated physiological adjustments in humans using multiple psychophysiological and brain measures. When participants were exposed to a dynamically approaching, uncontrollable threat, attentive freezing was augmented, as indicated by an increase in skin conductance, fear bradycardia, and potentiation of the startle reflex. In contrast, when participants had the opportunity to actively avoid the approaching threat, attention switched to response preparation, as indicated by an inhibition of the startle magnitude and by a sharp drop of the probe-elicited P3 component of the evoked brain potentials. These new findings on the dynamics of defensive behaviors form an important intersection between animal and human research and have important implications for understanding fear and anxiety-related disorders.
How acute stress may enhance subsequent memory for threat stimuli outside the focus of attention: DLPFC-amygdala decoupling
DOI:10.1016/j.neuroimage.2018.01.010
URL
PMID:29329979
[本文引用: 3]
Stress-related disorders, e.g., anxiety and depression, are characterized by decreased top-down control for distracting information, as well as a memory bias for threatening information. However, it is unclear how acute stress biases mnemonic encoding and leads to prioritized storage of threat-related information even if outside the focus of attention. In the current study, healthy adults (N = 53, all male) were randomly assigned to stress induction using the socially evaluated cold-pressor test (SECPT) or a control condition. Participants performed a task in which they were required to identify a target letter within a string of letters that were either identical to the target and thereby facilitating detection (low distractor load) or mixed with other letters to complicate the search (high load). Either a fearful or neutral face was presented on the background, outside the focus of attention. Twenty-four hours later, participants were asked to perform a surprise recognition memory test for those background faces. Stress induction resulted in increased cortisol and negative subjective mood ratings. Stress did not affect visual search performance, however, participants in the stress group showed stronger memory compared to the control group for fearful faces in the low attentional load condition. Critically, the stress induced memory bias was accompanied by decoupling between amygdala and DLFPC during encoding, which may represent a mechanism for decreased ability to filter task-irrelevant threatening background information. The current study provides a potential neural account for how stress can produce a negative memory bias for threatening information even if presented outside the focus of attention. Despite of an adaptive advantage for survival, such tendencies may ultimately also lead to generalized fear, a possibility requiring additional investigation.
Cognitive neuroscience of stress
Stress has profound impacts on physiology, the brain, cognition and behaviour. In response to stress, the brain initiates several neuromodulatory and endocrine systems. This leads to the release of stress-sensitive mediators, including neuromodulators, hormones and neuropeptides, which in turn feed back on the brain and thus alters cognition and behaviour. As the progress of sophisticated cognitive neuroimaging techniques, cognitive neuroscientists have begun to elucidate the psychological and neural mechanisms underlying detrimental and beneficial effects of stress on emotion, cognition and behaviour in the human brain. Stress-related neuromodulations occur at various stages of information processing, from the initial vigilance, attention, executive functions, learning and memory, and emotion. We reviewed recent advances in cognitive neuroscience of stress and proposed a multidimensional model of stress for better understanding how stress-sensitive neuromodulatory and endocrine systems interplay to impact the brain, cognition and behaviour in humans.
应激的认知神经科学研究
Individual differences in anxiety predict neural measures of visual working memory for untrustworthy faces
DOI:10.1093/scan/nst189
URL
[本文引用: 6]
When facing strangers, one of the first evaluations people perform is to implicitly assess their trustworthiness. However, the underlying processes supporting trustworthiness appraisal are poorly understood. We hypothesized that visual working memory (VWM) maintains online face representations that are sensitive to physical cues of trustworthiness, and that differences among individuals in representing untrustworthy faces are associated with individual differences in anxiety. Participants performed a change detection task that required encoding and maintaining for a short interval the identity of one face parametrically manipulated to be either trustworthy or untrustworthy. The sustained posterior contralateral negativity (SPCN), an event-related component (ERP) time-locked to the onset of the face, was used to index the resolution of face representations in VWM. Results revealed greater SPCN amplitudes for trustworthy faces when compared with untrustworthy faces, indicating that VWM is sensitive to physical cues of trustworthiness, even in the absence of explicit trustworthiness appraisal. In addition, differences in SPCN amplitude between trustworthy and untrustworthy faces correlated with participants' anxiety, indicating that healthy college students with sub-clinical high anxiety levels represented untrustworthy faces in greater detail compared with students with sub-clinical low anxiety levels. This pattern of findings is discussed in terms of the high flexibility of aversive/avoidance and appetitive/approach motivational systems.
A meta-analysis of the magnitude of biased attention in depression
DOI:10.1002/da.20755
URL
PMID:21049527
[本文引用: 1]
In this quantitative review, we examined the magnitude of attentional biases to negative stimuli in depression. Results from 29 empirical studies examining emotional Stroop or dot probe results in depressed participants (clinical depression, nonclinical dysphoria, and subjects undergoing depressive mood induction) were examined. Studies using the emotional Stroop task yielded marginally significant evidence of a difference between depressed and nondepressed samples, whereas those using the dot probe task showed significant differences between groups (d = 0.52). We found no evidence for significant moderation of these effects by age, sex, type of depressed sample, year of publication, stimulus presentation duration, or type of stimuli (verbal or nonverbal), although statistical power for these tests was limited. These results support the existence of biased attention to negative information in depression.
Acute psychological stress reduces working memory-related activity in the dorsolateral prefrontal cortex
DOI:10.1016/j.biopsych.2009.03.006
URL
PMID:19403118
[本文引用: 3]
Acute psychological stress impairs higher-order cognitive function such as working memory (WM). Similar impairments are seen in various psychiatric disorders that are associated with higher susceptibility to stress and with prefrontal cortical dysfunctions, suggesting that acute stress may play a potential role in such dysfunctions. However, it remains unknown whether acute stress has immediate effects on WM-related prefrontal activity.
The influence of stress on attentional bias to threat: An angry face and a noisy crowd
DOI:10.3758/s13423-018-1538-2
URL
PMID:30353494
[本文引用: 1]
During stress, attentional capture by threatening stimuli may be particularly adaptive. Individuals are more efficient at identifying threatening faces in a crowd than identifying nonthreatening faces (e.g., öhman et al., Journal of Experimental Psychology: General, 130(3): 466-478, 2001a, öhman et al., Journal of Personality and Social Psychology, 80(3): 381-396, 2001b). However, under conditions of stress, when attention to threat may be most critical, cognitive processes are generally disrupted. The present study explored the attentional advantage of threatening stimuli under stressful conditions. We exposed participants to either high or low stress conditions during a visual search task displaying threatening and nonthreatening facial targets among distractors. Participants' accuracy, reaction times, and self-reported stress were measured. Stress introduced a speed-accuracy trade-off: participants in the high-stress condition were faster, but less accurate, than participants in the low-stress condition. Although both groups of participants showed relative performance advantages in detecting threatening compared with nonthreatening stimuli, this advantage was markedly larger for participants in the high-stress condition. This suggests that the established stress-mediated increase in the activity of the ventral neural network responsible for the reorienting of attention may have enhanced the ability to detect threatening stimuli or buffered the disruptive effects of stress on this process. Our findings highlight the potentially adaptive nature of stress disruption on attentional processes and align research on the anger superiority effect and automated attentional processes under stress.
Selective attention and threat: Quick orienting versus slow disengagement and two versions of the dot probe task
DOI:10.1016/j.brat.2006.04.004
URL
[本文引用: 2]
AbstractThe dot probe task is often used to assess attentional bias in anxiety, but some aspects need clarification. First, the results, which are traditionally summarized in an attentional bias index, do not allow for distinguishing between different selective attention processes; orienting and disengagement. Second, different versions of the dot probe task have been used with unknown relative merits. Participants (N=133) completed two versions of the dot probe task: the detection task (i.e. is there a probe?) and the differentiation task (i.e. what sort of probe is it?). The analysis carried out allowed for a differentiation between orienting and disengagement. The main finding was that trait anxiety is related to disengagement difficulties and not to speed of orienting. Concerning the relative merits of the two dot probe task versions, the results suggest that the detection task may be superior to the differentiation task. Implications for past and future research are discussed.]]>
The influence of acute stress on attention mechanisms and its electrophysiological correlates
DOI:10.3389/fnbeh.2014.00353
URL
PMID:25346669
[本文引用: 3]
FOR THE SELECTION OF RELEVANT INFORMATION OUT OF A CONTINUOUS STREAM OF INFORMATION, WHICH IS A COMMON DEFINITION OF ATTENTION, TWO CORE MECHANISMS ARE ASSUMED: a competition-based comparison of the neuronal activity in sensory areas and the top-down modulation of this competition by frontal executive control functions. Those control functions are thought to bias the processing of information toward the intended goals. Acute stress is thought to impair these frontal functions through the release of cortisol. In the present study, subjects had to detect a luminance change of a stimulus and ignore more salient but task irrelevant orientation changes. Before the execution of this task, subjects underwent a socially evaluated cold pressor test (SECPT) or a non-stressful control situation. The SECPT revealed reliable stress response with a significant increase of cortisol and alpha-amylase. Stressed subjects showed higher error rates than controls, particularly in conditions which require top-down control processing to bias the less salient target feature against the more salient and spatially separated distracter. By means of the EEG, subjects who got stressed showed a reduced allocation to the relevant luminance change apparent in a modulation of the N1pc. The following N2pc, which reflects a re-allocation of attentional resources, supports the error pattern. There was only an N2pc in conditions, which required to bias the less salient luminance change. Moreover, this N2pc was decreased as a consequence of the induced stress. These results allow the conclusion that acute stress impairs the intention-based attentional allocation and enhances the stimulus-driven selection, leading to a strong distractibility during attentional information selection.
A common neural mechanism for preventing and terminating the allocation of attention
DOI:10.1523/JNEUROSCI.1864-12.2012
URL
[本文引用: 2]
Much is known about the mechanisms by which attention is focused to facilitate perception, but little is known about what happens to attention after perception of the attended object is complete. One possibility is that the focus of attention passively fades. A second possibility is that attention is actively terminated after the completion of perception so that the brain can be prepared for the next target. The present study investigated this issue with event-related potentials in humans, focusing on the N2pc component (a neural measure of attentional deployment) and the Pd component (a neural measure of attentional suppression). We found that active suppression occurred both to prevent the allocation of attention to known distractors and to terminate attention after the perception of an attended object was complete. In addition, the neural measure of active suppression was correlated with a behavioral measure of trial-to-trial variations in the allocation of attention. Active suppression therefore appears to be a general-purpose mechanism that both prevents and terminates the allocation of attention.
HPA axis activation by a socially evaluated cold-pressor test
DOI:10.1016/j.psyneuen.2008.03.001
URL
[本文引用: 2]
SummaryThe cold-pressor test (CPT) in which subjects immerse their hand in ice water is among the most commonly used laboratory stressors. While the CPT elicits strong sympathetic nervous system activation, cortisol elevations indicative for the reactivity of the hypothalamus–pituitary–adrenal (HPA) axis are moderate to low in response to the CPT. In the present study, we assessed whether cortisol responses to the CPT can be increased by adding social-evaluative elements. Therefore, 70 healthy young men immersed their hand in ice or warm water and were watched by a woman and videotaped during hand immersion or not. While the standard CPT and the socially evaluated cold-pressor test (SECPT) led to comparable increases in blood pressure and subjective stress ratings, saliva cortisol elevations and the proportion of subjects showing a saliva cortisol response (defined as increase >2 nmol/l) were significantly higher after the SECPT. Social evaluation during hand immersion in warm water did not affect saliva cortisol levels suggesting that both social evaluation and a challenge are required for HPA axis activation. These findings indicate that the incorporation of social-evaluative elements increases HPA axis responses to the CPT. The SECPT can serve as a tool for future stress research.]]>
Stress-induced enhancement of response inhibition depends on mineralocorticoid receptor activation
DOI:10.1016/j.psyneuen.2013.05.001
URL
[本文引用: 1]
Stress is a well-known modulator of cognitive functions. These effects are, at least in part, mediated by glucocorticoid stress hormones which act via two receptor types in the brain, glucocorticoid receptors (GR) and mineralocorticoid receptors (MR). Here, we examined whether stress affects inhibitory control processes and, if so, whether these effects are mediated by the MR. To this end, healthy participants received 300 mg of the MR antagonist spironolactone or a placebo and underwent a stressor (socially evaluated cold pressor test) or a non-stressful control task 90 min later. Shortly after the stressor, participants performed a stop-signal task that required them to rapidly suppress a well-established response whenever a tone was presented. Results revealed that stress enhanced response inhibition in the stop-signal task and that this enhancement was abolished by spironolactone. Our results show that stress may facilitate inhibitory control and that these effects depend on MR functioning. (C) 2013 Elsevier Ltd.
Stress effects on memory: An update and integration
DOI:10.1016/j.neubiorev.2011.07.002
URL
[本文引用: 1]
It is well known that stressful experiences may affect learning and memory processes. Less clear is the exact nature of these stress effects on memory: both enhancing and impairing effects have been reported. These opposite effects may be explained if the different time courses of stress hormone, in particular catecholamine and glucocorticoid, actions are taken into account. Integrating two popular models, we argue here that rapid catecholamine and non-genomic glucocorticoid actions interact in the basolateral amygdala to shift the organism into a 'memory formation mode' that facilitates the consolidation of stressful experiences into long-term memory. The undisturbed consolidation of these experiences is then promoted by genomic glucocorticoid actions that induce a 'memory storage mode', which suppresses competing cognitive processes and thus reduces interference by unrelated material. Highlighting some current trends in the field, we further argue that stress affects learning and memory processes beyond the basolateral amygdala and hippocampus and that stress may pre-program subsequent memory performance when it is experienced during critical periods of brain development. (C) 2011 Elsevier Ltd.
Ten years of research with the Socially Evaluated Cold Pressor Test: Data from the past and guidelines for the future
DOI:10.1016/j.psyneuen.2018.03.010
URL
PMID:29573884
[本文引用: 2]
Ten years ago, the Socially Evaluated Cold Pressor Test (SECPT) was introduced as a standardized protocol for the efficient experimental stress induction in humans. In short, the 3 min SECPT, which can be conducted by only a single experimenter, combines a physiological challenge (hand immersion into ice water) with socio-evaluative elements. The purpose of this article is twofold. First, we aim to evaluate the subjective and physiological responses elicited by the SECPT. To this end, we pooled data from 21 studies from our lab and systematically analyzed the response profile to the SECPT. Our analyses show that the SECPT leads, both in men and women, to striking increases in subjective stress levels, autonomic arousal, and cortisol, albeit the cortisol response is typically somewhat less pronounced than in the Trier Social Stress Test. Second, we aim to provide guidelines for conducting the SECPT, in order to foster homogenization of the SECPT procedure across (and within) labs. In sum, we argue that the SECPT is a highly efficient tool to induce stress and activate major stress systems in a laboratory context, in particular if the guidelines that we outline here are followed.
Interhemispheric ERP asymmetries over inferior parietal cortex reveal differential visual working memory maintenance for fearful versus neutral facial identities
DOI:10.1111/j.1469-8986.2010.01046.x
URL
[本文引用: 1]
The goal of the present investigation was to discover whether visual working memory maintenance for faces is modulated by facial expression using event-related potentials (ERPs). Each trial consisted of two sequential arrays, a memory array and a test array, each including either two or four faces with neutral or fearful expressions. The faces were displayed to the left and to the right of a central fixation cross. Two central arrows cued participants to encode one face or two faces displayed on one side of the memory array. The sustained posterior contralateral negativity (SPCN) component of the ERP time-locked to the onset of the memory array was used as an index of visual working memory maintenance. Visual working memory performance was quantified using indexes of memory capacity (Cowan's K and K-iterative), a standard index of sensitivity (d'), and reaction times (RTs). Relative to neutral faces, superior memory and longer change-detection RTs to fearful face identities were observed when two faces were displayed on the cued side of the memory array. Fearful faces elicited an enhanced SPCN relative to neutral faces, especially when only one face was displayed on the cued side of the memory array. These findings suggest increased maintenance in visual working memory of faces with a fearful expression relative to faces with a neutral expression and that the representational format in which fearful faces are stored in memory may be characterized by enhanced resolution relative to that subtended in the maintenance of neutral faces.
Stress potentiates early and attenuates late stages of visual processing
DOI:10.1523/JNEUROSCI.3384-10.2011
URL
[本文引用: 1]
Stress can fundamentally alter neural responses to incoming information. Recent research suggests that stress and anxiety shift the balance of attention away from a task-directed mode, governed by prefrontal cortex, to a sensory-vigilance mode, governed by the amygdala and other threat-sensitive regions. A key untested prediction of this framework is that stress exerts dissociable effects on different stages of information processing. This study exploited the temporal resolution afforded by event-related potentials to disentangle the impact of stress on vigilance, indexed by early perceptual activity, from its impact on task-directed cognition, indexed by later postperceptual activity in humans. Results indicated that threat of shock amplified stress, measured using retrospective ratings and concurrent facial electromyography. Stress also double-dissociated early sensory-specific processing from later task-directed processing of emotionally neutral stimuli: stress amplified N1 (184-236 ms) and attenuated P3 (316-488 ms) activity. This demonstrates that stress can have strikingly different consequences at different processing stages. Consistent with recent suggestions, stress amplified earlier extrastriate activity in a manner consistent with vigilance for threat (N1), but disrupted later activity associated with the evaluation of task-relevant information (P3). These results provide a novel basis for understanding how stress can modulate information processing in everyday life and stress-sensitive disorders.
STAI manual for the state-trait anxiety inventory
DOI:10.1177/1120700019889334
URL
PMID:31838874
[本文引用: 2]
Robotic-arm assisted surgery aims to reduce manual errors and improve the accuracy of implant positioning and orientation during total hip arthroplasty (THA). The objective of this study was to assess the surgical team's learning curve for robotic-arm assisted acetabular cup positioning during THA.
Attentional selection and suppression in children and adults
DOI:10.1111/desc.12684
URL
PMID:29761932
[本文引用: 2]
D ). EEG data were collected from healthy young adults and typically developing children (9-15 years old) as they searched for a shape singleton target in either the absence or the presence of a salient-but-irrelevant color singleton distractor. The ERP results showed that a lateral shape target elicited a smaller N2pc in children compared with adults regardless of whether a distractor was present or not. Moreover, the target-elicited N2pc was always followed by a similar positivity in both age groups. Counterintuitively, a lateral salient-but-irrelevant distractor elicited a large PD in children with low behavioral accuracy, whereas high-accuracy children exhibited a small and "adult-like" PD . More importantly, we found no evidence for a correlation between the target-elicited N2pc and the distractor-elicited PD in either age group. Our results provide neurophysiological evidence for the developmental differences between target selection and distractor suppression. Compared with adults, 9-15-year-old children deploy insufficient attentional selection resources to targets but use "adult-like" or even more attentional suppression resources to resist irrelevant distractors. A video abstract of this article can be viewed at: https://www.youtube.com/watch?v=NhWapx0d75I.]]>
Attentional control moderates the relationship between social anxiety symptoms and attentional disengagement from threatening information
DOI:10.1016/j.jbtep.2015.05.008
URL
PMID:26072705
[本文引用: 2]
Social anxiety is characterized by biased attentional processing of social information. However, heterogeneity of extant findings suggests that it may be informative to elucidate individual difference factors that modulate the processing of emotional information. The current study examined whether individual differences in components of attentional control (AC--shifting and focusing) moderated the link between social anxiety and attentional engagement and disengagement biases for threat-relevant cues.
Neural regulation of endocrine and autonomic stress responses
DOI:10.1038/nrn2647
URL
PMID:19469025
[本文引用: 1]
The survival and well-being of all species requires appropriate physiological responses to environmental and homeostatic challenges. The re- establishment and maintenance of homeostasis entails the coordinated activation and control of neuroendocrine and autonomic stress systems. These collective stress responses are mediated by largely overlapping circuits in the limbic forebrain, the hypothalamus and the brainstem, so that the respective contributions of the neuroendocrine and autonomic systems are tuned in accordance with stressor modality and intensity. Limbic regions that are responsible for regulating stress responses intersect with circuits that are responsible for memory and reward, providing a means to tailor the stress response with respect to prior experience and anticipated outcomes.
From specificity to sensitivity: How acute stress affects amygdala processing of biologically salient stimuli
DOI:10.1016/j.biopsych.2009.05.014
URL
PMID:19596123
[本文引用: 1]
A vital component of an organism's response to acute stress is a surge in vigilance that serves to optimize the detection and assessment of threats to its homeostasis. The amygdala is thought to regulate this process, but in humans, acute stress and amygdala function have up to now only been studied in isolation. Hence, we developed an integrated design using functional magnetic resonance imaging to investigate the immediate effects of controlled stress induction on amygdala function.
Attentional selection and suppression in children with attention-deficit/hyperactivity disorder
DOI:10.1016/j.bpsc.2016.01.004
URL
PMID:29560866
[本文引用: 2]
D]).]]>
Standardization and assessment of college students' facial expression of emotion
DOI:10.1002/1097-4679(195710)13:4&lt;396::aid-jclp2270130421&gt;3.0.co;2-e URL PMID:13463148 [本文引用: 1]
大学生面孔表情材料的标准化及其评定
Effects of acute psychosocial stress on working memory related brain activity in men
DOI:10.1002/hbm.20945
URL
PMID:20127868
[本文引用: 1]
Acute psychosocial stress in humans triggers the release of glucocorticoids (GCs) and influences performance in declarative and working memory (WM) tasks. These memory systems rely on the hippocampus and prefrontal cortex (PFC), where GC-binding receptors are present. Previous studies revealed contradictory results regarding effects of acute stress on WM-related brain activity. We combined functional magnetic resonance imaging with a standardized psychosocial stress protocol to investigate the effects of acute mental stress on brain activity during encoding, maintenance, and retrieval of WM. Participants (41 healthy young men) underwent either a stress or a control procedure before performing a WM task. Stress increased salivary cortisol levels and tended to increase WM accuracy. Neurally, stress-induced increases in cortical activity were evident in PFC and posterior parietal cortex (PPC) during WM maintenance. Furthermore, hippocampal activity was modulated by stress during encoding and retrieval with increases in the right anterior hippocampus during WM encoding and decreases in the left posterior hippocampus during retrieval. Our study demonstrates that stress increases activity in PFC and PPC specifically during maintenance of items in WM, whereas effects on hippocampal activity are restricted to encoding and retrieval. The finding that psychosocial stress can increase and decrease activity in two different hippocampal areas may be relevant for understanding the often-reported phase-dependent opposing behavioral effects of stress on long-term memory.
Stress sensitizes the brain: Increased processing of unpleasant pictures after exposure to acute stress
A key component of acute stress is a surge in vigilance that enables a prioritized processing of highly salient information to promote the organism's survival. In this study, we investigated the neural effects of acute stress on emotional picture processing. ERPs were measured during a deep encoding task, in which 40 male participants categorized 50 unpleasant and 50 neutral pictures according to arousal and valence. Before picture encoding, participants were subjected either to the Socially Evaluated Cold Pressor Test (SECPT) or to a warm water control procedure. The exposure to the SECPT resulted in increased subjective and autonomic (heart rate and blood pressure) stress responses relative to the control condition. Viewing of unpleasant relative to neutral pictures evoked enhanced late positive potentials (LPPs) over centro-parietal scalp sites around 400 msec after picture onset. Prior exposure to acute stress selectively increased the LPPs for unpleasant pictures. Moreover, the LPP magnitude for unpleasant pictures following the SECPT was positively associated with incidental free recall performance 24 hr later. The present results suggest that acute stress sensitizes the brain for increased processing of cues in the environment, particularly priming the processing of unpleasant cues. This increased processing is related to later long-term memory performance.
Long-term academic stress increases the late component of error processing: An ERP study
DOI:10.1016/j.biopsycho.2014.03.002
URL
[本文引用: 1]
Exposure to long-term stress has a variety of consequences on the brain and cognition. Few studies have examined the influence of long-term stress on event related potential (ERP) indices of error processing. The current study investigated how long-term academic stress modulates the error related negativity (Ne or ERN) and the error positivity (Pe) components of error processing. Forty-one male participants undergoing preparation for a major academic examination and 20 non-exam participants completed a Go-NoGo task while ERP measures were collected. The exam group reported higher perceived stress levels and showed increased Pe amplitude compared with the non-exam group. Participants' rating of the importance of the exam was positively associated with the amplitude of Pe, but these effects were not found for the Ne/ERN. These results suggest that long-term academic stress leads to greater motivational assessment of and higher emotional response to errors. (C) 2014 Elsevier B.V.
Attentional bias towards threat: Facilitated attentional orienting or impaired attentional disengagement?
DOI:10.3724/SP.J.1042.2014.01129
URL
[本文引用: 1]
attentional bias towards threat is a common phenomenon in anxious as well as healthy individuals. There are two potential cognitive mechanisms of attentional bias. The facilitated attentional orienting is known as threat stimuli are detected faster than neutral stimuli. The impaired attentional disengagement is defined as harder to disengage attention from threat stimuli compare to neutral stimuli. The amygdala- anterior cingulate cortex network is the potential neural base of facilitated attentional orienting, while the orbitofrontal cortex is the key area of attentional disengagement. The properties of threat stimuli and characteristic of participants are main factors which affect attentional bias. The future studies should focus on the relationship between attentional orienting and attentional disengagement as well as the neural base of attentional bias.
对威胁刺激的注意偏向: 注意定向加速还是注意解除困难?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}