

1 引言
从古至今, “不患寡而患不均”的思想始终促使着人们不断追求公平(吴燕, 周晓林, 2012)。公平作为一种基本的社会准则, 在人类生活中起着重要的作用, 它不仅保障了个体的基本权利, 同时也维护了社会的稳定发展。大量研究显示, 当个体面对不公平时, 个体的行为反应具有跨文化的一致性(Knoch, Pascual-Leone, Meyer, Treyer, & Fehr, 2006), 具体表现为牺牲自我利益以惩罚该种行为, 而惩罚被证明是制裁违反公平规范的有效方法, 有利于人际公平的形成(Fehr & Gächter, 2000; Henrich et al., 2006; Makwana, Grön, Fehr, & Hare, 2015; Wu et al., 2014)。
目前对人际公平的研究主要采用行为经济学的博弈范式, 其中最后通牒博弈任务(Ultimatum Game, UG) (Güth, Schmittberger, & Schwarze, 1982)是应用较为广泛的一种实验范式(Feng, Luo, & Krueger, 2015; Gabay, Radua, Kempton, & Mehta, 2014)。经典的最后通牒博弈任务包含两个角色, 提议者(proposer)和接受者(recipient)。要求他们对一笔金钱进行分配, 先由提议者进行分配, 然后接受者决定是否接受当前的分配方案。如果接受者认可该分配方案, 就按照该提议进行分配; 如果接受者拒绝, 则双方均无任何收益。Spitzer, Fischbacher, Herrnberger, Grön和Fehr (2007)采用修订版的最后通牒博弈任务探究了惩罚对于公平规范遵从的作用。惩罚条件下, 若接受者认为分配方案不公平, 可以采取惩罚措施, 即接受者每拿出1元, 就相应减少分配者5元。研究结果显示, 无惩罚下提议者的分配金额仅为总金额的10%, 而惩罚下分配金额达到总金额的40%, 更接近平均分配(50%)。另外, 惩罚条件下, 分配者的分配越不公平(与50的差值越大), 接受者的惩罚金额越大。由此可见, 惩罚在促进人际公平方面起着重要作用(Makwana et al., 2015)。
随着脑成像技术的日益成熟, 研究者们利用功能性磁共振成像技术(fMRI)深入探究最后通牒博弈任务中提议者和接受者遵从公平规范的神经机制。已有研究显示, 相比无惩罚条件, 惩罚条件下提议者的分配金额更高, 分配过程中双侧背外侧前额叶皮层(DLPFC)、腹外侧前额叶皮层(VLPFC)、眶额皮层(OFC)以及双侧尾状核(caudate nucleus)的激活程度增强, 惩罚条件下右侧背外侧前额叶皮层、左侧眶额皮层、右侧尾状核激活程度的增强与公平规范遵从程度的提升(惩罚下分配金额减去无惩罚下分配金额)呈正相关(Spitzer et al., 2007)。这表明遵守公平规范可以激活与认知控制(Miller & Cohen, 2001)、优势反应(自私自利)抑制(Aron, Robbins, & Poldrack, 2004)、惩罚威胁评估(O'Doherty, Kringelbach, Rolls, Hornak, & Andrews, 2001)以及不确定奖赏与惩罚刺激加工相关的脑区(Delgado, Locke, Stenger, & Fiez, 2003)。同时, 研究者还发现公平规范遵从的神经通路具有偏侧化的特点, 使用经颅直流电刺激(tDCS)增强右侧DLPFC的激活程度可显著提升提议者的分配金额, 使其更接近平均分配; 而抑制该脑区则会降低分配方案的公平性(Ruff, Ugazio, & Fehr, 2013)。
对接受者遵从公平规范脑机制的研究显示, 存在两个交互作用的神经系统支持这一行为, 包括反射性直觉系统(reflexive and intuitive system)和反省深思系统(reflective and deliberate system)。前者由前脑岛(AI)、杏仁核(amygdala)和腹内侧前额叶皮层(VMPFC)组成, 涉及识别及评价社会规范违反行为, 产生惩罚动机。后者由背侧前扣带回(dACC)、背外侧前额叶皮层(DLPFC)、腹外侧前额叶皮层(VLPFC)和背内侧前额叶皮层(DMPFC)组成, 参与重评及冲突的调节, 抑制自利行为, 执行惩罚措施(Feng et al., 2015)。研究者们同样采用了无创的干预技术探究了接受者在遵从公平规范中DLPFC功能的偏侧化, 发现当右侧DLPFC被抑制时, 接受者面对不公平分配时惩罚行为显著减少(Baumgartner, Knoch, Hotz, Eisenegger, & Fehr, 2011; Knoch et al., 2008; Knoch et al., 2006)。以上研究结果均提示右侧DLPFC在经济博弈中可能负责自利反应的抑制, 让个体更好地遵从公平规范, 一定程度上也暗示了公平规范遵从下提议者和接受者右侧DLPFC神经活动的相似性。
过往研究主要集中于对经济博弈中某一方决策行为和神经机制的探讨, 但是人际公平的形成可能是互动双方重复博弈的结果(Spitzer et al., 2007)。而重复的博弈是一种互为因果的过程, 个体的行为结果既是一种反馈, 也是其互动对象行为的前提(Shaw et al., 2018)。因此只考察单个个体的大脑活动, 并不足以揭示由互动双方共同完成的社会认知活动的脑机制。所以本研究中我们从单脑(single- brain)转向多脑(multi-brain) (Hasson, Ghazanfar, Galantucci, Garrod, & Keysers, 2012), 通过分析脑-脑互动的变化规律, 从群体脑水平上揭示人际公平形成的神经机制。超扫描技术(hyperscanning)为人际互动下跨脑机制的探究提供了新途径(Montague et al., 2002), 通过对互动的多个脑活动进行同步记录, 并分析脑间活动同步性(Interpersonal Neural Synchronization, INS), 从群体角度上提供了社会互动脑机制的新机制。该技术已在社会认知神经科学的多个领域有所应用, 研究涉及合作竞争(Cui, Bryant, & Reiss, 2012; Pan, Cheng, Zhang, Li, & Hu, 2017)、行为模仿(Holper, Scholkmann, & Wolf, 2012; Pan, Novembre, Song, Li, & Hu, 2018)、师生互动(Zheng et al., 2018)等。这些研究一致表明, 社会认知相关脑区, 特别是与“心理理论”相关脑区的INS是衡量人际互动重要的神经指标。在一项金钱分配游戏中, 相比隔板遮挡的情境, 面对面互动下双方右侧颞-顶联合区(TPJ)的INS增强。这可能是因为面对面的交流提供了外显的社会线索, 例如面部表情、肢体动作等, 使得双方能够更好地揣测对方心理, 达成共享意图(Tang et al., 2016)。Liu, Saito和Oi (2015)采用轮流下棋的游戏探究了合作与竞争的神经机制。任务中包含两个角色, 创建者(builder)和搭档(partner)。合作任务中, 搭档需要辅助创建者复制给定的棋局。竞争任务中, 搭档要尽可能阻碍创建者对棋局的复制。双方轮流摆放棋子, 每人摆放四次。研究者发现, 竞争任务中右侧额下回(IFG)出现了INS, 但是这一现象并不存在合作任务中。研究者认为竞争任务中, 双方的卷入度更高, 更加积极地推断他人的心理意图并调整自己的行为策略。由此可见, 互动双方在相互博弈的过程中, “心理理论”相关脑区的交互性更强。
相比其他的脑功能成像技术, 功能近红外光谱技术(functional near-infrared spectroscopy, fNIRS)在成本、对头动的容忍性以及研究的生态效度等方面具有优势(Scholkmann, Holper, Wolf, & Wolf, 2013)。因此, 本研究将结合修改版的最后通牒博弈任务与基于fNIRS的超扫描技术, 设置惩罚和无惩罚两个条件, 分析不同条件下分配金额和脑间活动同步性的差异, 从互动视角下揭示人际公平形成的脑机制。基于以往研究结果, 惩罚可以促使个体在经济决策中遵从公平规范, 我们预期(1)惩罚下分配金额高于无惩罚条件, 且更趋近平均分配。与此同时, 遵从公平规范需要个体抑制优势反应(自利行为), 且该过程中个体间的交互性更强。因此, 我们预期(2)相比无惩罚条件, 惩罚下提议者和接受者右侧背外侧前额叶以及“心理理论”相关脑区的INS更强。
2 方法
2.1 被试
共招募大学在读本科生44名(M = 21.3岁, SD = 1.6岁), 其中男生26人, 女生18人, 共形成22对同性别被试对, 每组被试在实验前互不认识。所有被试均为右利手, 无脑部疾病或者精神疾病, 视力或者矫正视力均正常。实验前, 所有被试签署了知情同意。实验后, 根据被试在实验过程中的实际表现给与相应的被试费。
2.2 实验任务
实验采用Spitzer等人(2007)基于最后通牒博弈任务修改而成的金钱分配任务, 要求两名被试完成多轮金钱分配, 金额为100元。实验一共分为两个条件, 惩罚条件和无惩罚条件。惩罚条件下, 由提议者(A)提出分配方案。如果接受者(B)认可分配方案, 就按照该方案进行金钱分配。如果B认为分配方案不合理, 可以通过减少自己拥有的金钱来对提议者进行惩罚, 惩罚比率为1:5, 即接受者每拿出1元, 就相应地减少提议者5元。因此, 为了收益最大化, A应适当调整分配方案, 找到B所能接受的最小分配金额并努力维持; 而B应通过惩罚的反馈方式, 找到A所能给与的最大分配金额并尽力保持。无惩罚条件下, 无论B是否认可A的分配方案, 都只能无条件接受。
2.3 实验流程
2名被试进入实验室后, 面对面而坐, 中间有隔板遮挡(如图1A所示)。实验开始前, 被试双方通过抽签的方式来决定任务角色(提议者A和接受者B), 任务中角色固定。每一轮试次中, 两人都先获得25元, 这保证了惩罚条件下, 即便A分配给B 0元, B也有资本对A进行惩罚。之后, A对100元进行分配。试次始于3~6 s的注视点, 接着呈现线索2 s, 5 : 1代表惩罚条件, 0:0代表无惩罚条件。任务线索呈现完毕后, A思考分配方案2 s, 而后提出分配方案, 通过按“←”、“→”键控制绿色光标的移动, 从而增加或者减少分配金额, 时间不限, 分配完后A按“↓”键进行确认。而后呈现该分配方案2 s, 之后B进行反馈。在5 : 1的线索下, B既可以接受, 也可以通过减少25元中的部分或全部金钱来减少A相应的5倍金钱。通过按“1”、“3”键控制绿色光标的移动, B可以增加或减少惩罚金额, 惩罚完后按“2”键进行确认。需要注意的是, 惩罚过程中, 接受者仅能减少提议者的金额直至为0元。在0 : 0的线索下, B只能按“2”键接受A的分配方案。最后向被试双方呈现分配结果2 s。任务中A和B电脑屏幕上所呈现的内容一致, 即B可以实时看见A的分配情况, A也能实时获取B的反馈情况(如图1B所示)。
图1
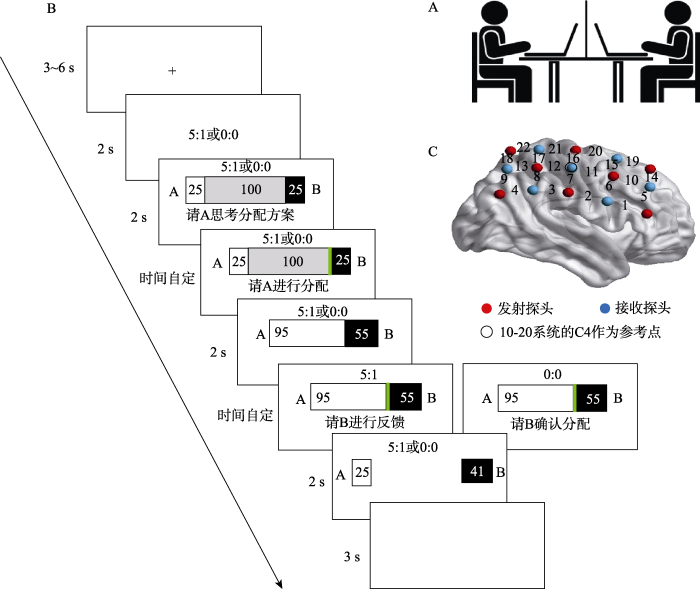
任务共包含50个试次, 惩罚条件和无惩罚条件各25个试次, 两种条件的试次以伪随机的方式进行。任务开始前, 被试静息40 s。完成25个试次后, 被试休息2分钟, 然后完成剩余的25个试次。实验前告知被试, 被试费由基本被试费(完成实验)和任务过程中积累金额的一定百分比组成。
2.4 数据收集
通过E-prime 2.0软件呈现任务刺激并采集行为学数据。另外, 采用Hitachi (日立) ETG-4000来记录被试在实验任务中大脑氧合血红蛋白(oxy- hemoglobin, Hbo)以及脱氧血红蛋白(deoxy- hemoglobin, Hbr)浓度的变化。将一块3×5的探头板置于泳帽中, 放置在每名被试的大脑右侧。根据国际标准10-20系统, 将探头板正中间的探测器放置在C4的位置, 探头板沿着矢状参考曲线进行放置(如图1C所示), 覆盖脑区为额叶和顶叶。每个探头板由8个发射器和7个探测器组成, 探头之间间隔3cm, 每个探头板有22个通道, 每个通道的具体位置参考Jichi University提供的定位模板(http://www.jichi.ac.jp/brainlab/virtual_registration/Result3x5_E.html, 如表1所示) (Reindl, Gerloff, Scharke, & Konrad, 2018)。采样频率为10 Hz。
表1 通道位置
| 通道 | MNI坐标 | AAL分区 | 布鲁德曼分区 | ||||
|---|---|---|---|---|---|---|---|
| x | y | z | 脑区 | 概率 | 脑区 | 概率 | |
| 1 | 59.49 | 21.37 | 20.43 | 右侧三角部额下回 | 0.67 | 44-岛盖部, 布洛卡区部分 | 0.53 |
| 2 | 66.87 | -7.18 | 29.10 | 右侧中央后回 | 1.00 | 43-中央下区 | 1.00 |
| 3 | 67.53 | -33.40 | 34.90 | 右侧缘上回 | 1.00 | 40-缘上回, 威尔尼克区部分 | 0.72 |
| 4 | 59.42 | -59.03 | 33.70 | 右侧角回 | 1.00 | 39-角回, 威尔尼克区部分 | 0.97 |
| 5 | 50.69 | 36.73 | 27.05 | 右侧额中回 | 0.52 | 45-布洛卡区三角区 | 1.00 |
| 6 | 58.80 | 7.96 | 36.56 | 右侧中央前回 | 0.97 | 6-前运动皮层和辅助运动皮层 | 0.78 |
| 7 | 63.88 | -20.28 | 43.97 | 右侧缘上回 | 0.76 | 1-初级躯体感觉皮质 | 0.83 |
| 8 | 60.37 | -45.14 | 46.84 | 右侧顶下小叶 | 0.60 | 40-缘上回, 威尔尼克区部分 | 1.00 |
| 9 | 48.36 | -68.79 | 43.83 | 右侧角回 | 1.00 | 39-角回, 威尔尼克区部分 | 1.00 |
| 10 | 47.43 | 24.55 | 43.41 | 右侧额中回 | 0.99 | 44-岛盖部, 布洛卡区部分 | 0.51 |
| 11 | 54.49 | -4.25 | 50.69 | 右侧中央前回 | 0.55 | 6-前运动皮层和辅助运动皮层 | 0.97 |
| 12 | 56.89 | -31.30 | 54.81 | 右侧顶下小叶 | 0.72 | 1-初级躯体感觉皮质 | 0.68 |
| 13 | 49.68 | -55.49 | 55.37 | 右侧顶下小叶 | 0.84 | 40-缘上回, 威尔尼克区部分 | 0.81 |
| 14 | 33.83 | 37.74 | 46.08 | 右侧额中回 | 0.98 | 9-背外侧前额叶皮层 | 1.00 |
| 15 | 42.48 | 13.34 | 56.74 | 右侧额中回 | 1.00 | 9-背外侧前额叶皮层 | 0.79 |
| 16 | 47.64 | -17.53 | 62.57 | 右侧中央前回 | 0.52 | 4-初级运动皮层 | 0.65 |
| 17 | 43.34 | -42.84 | 63.31 | 右侧顶上小叶 | 0.55 | 2-初级躯体感觉皮质 | 0.52 |
| 18 | 32.38 | -63.83 | 61.62 | 右侧顶上小叶 | 0.99 | 7-躯体感觉联合皮层 | 1.00 |
| 19 | 27.55 | 24.62 | 58.16 | 右侧额上回 | 0.78 | 8-前额眼动区 | 0.99 |
| 20 | 34.22 | -1.43 | 65.56 | 右侧额上回 | 0.66 | 6-前运动皮层和辅助运动皮层 | 1.00 |
| 21 | 35.98 | -29.29 | 70.73 | 右侧中央后回 | 0.64 | 4-初级运动皮层 | 0.84 |
| 22 | 28.73 | -53.67 | 70.81 | 右侧顶上小叶 | 1.00 | 7-躯体感觉联合皮层 | 1.00 |
2.5 数据分析
2.5.1 行为数据
计算惩罚和无惩罚条件下每一位提议者A对接受者B的平均分配金额、A和B不同条件下的平均收益(包括分配前的初始资金25元), 并使用配对样本t检验比较不同实验条件下分配金额的差异以及A和B平均收益的差异。t检验的效应量为Cohen’s d, 0.2、0.5、0.8分别是小、中、大效应量。另外, 计算惩罚条件下接受者B对提议者A不公平分配(分配金额小于50元)的惩罚力度:
\[\frac {5×B对A的惩罚金额}{50×不公平试次数量-A对B不公平分配的总金额}\]
[分子:不公平试次(即分配金额小于50元)的总惩罚金额; 分母:不公平试次中提议者亏欠接受者的总金额。比值越大, 说明惩罚力度越大], 并使用皮尔逊相关计算惩罚力度与惩罚条件下分配金额的相关性。
2.5.2 fNIRS数据
在Matlab 2014a (Mathworks Inc., Natick, MA, USA)的操作环境下, 使用NIRS_SPM对数据进行预处理。采用血液动力响应函数(Hemodynamic Response Function, HRF)和离散余弦变换, 截止频率128 s (Discrete Cosine Transform, DCT)进行低通滤波和高通滤波, 从而消除仪器产生的噪音和呼吸心跳等引起的生理噪音(Ye, Tak, Jang, Jung, & Jang, 2009)。
鉴于氧合血红蛋白对任务刺激更加敏感(Hoshi, 2003), 在本研究中, 仅将其纳入进一步的数据分析。使用小波变换相干(Wavelet Transform Coherence, WTC)计算任务期间(线索阶段到反馈阶段)的脑间活动同步性(Interpersonal Neural Synchronization, INS) (Cui et al., 2012; Grinsted, Moore, & Jevrejeva, 2004)。首先, 对时间序列x (n)进行连续小波变换(Continuous Wavelet Transform,
CWT):\(W_{n}^{X}(s)=\sqrt{\frac{\delta t}{s}}\sum\limits_{{n}'=1}^{N}{x{n}'{{\psi }_{0}}}\left[ ({n}'-n)\frac{\delta t}{s} \right]\) (n:时间点, s:小波尺度, δt:采样周期(例如0.1 s), N:时间序列长度)。然后, 对两个时间序列x (n)和y (n)进行交叉小波变换(Cross Wavelet Transform, XWT):\({{W}^{XY}}(n,s)={{W}^{X}}(n,s){{W}^{Y*}}(n,s),\) *为复共轭。最后, 计算两个时间序列的小波变换相干性, \(R_{n}^{2}(s)=\frac{|S({{s}^{(-1)}}W{{_{n}^{X}}^{Y}}(s)){{|}^{2}}}{|S({{s}^{(-1)}}W_{n}^{X}(s)){{|}^{2}}\text{.}|S({{s}^{(-1)}}W_{n}^{Y}(s)){{|}^{2}}}\), S为平滑算子。
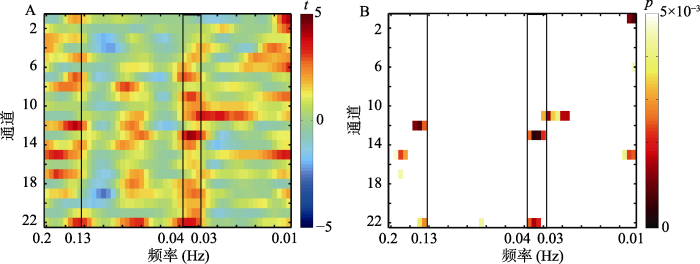
为了确保所选频段与任务相关, 基于以往超扫描研究(Nozawa, Sasaki, Sakaki, Yokoyama, & Kawashima, 2016; Pan et al., 2018; Zheng et al., 2018),将0.01~0.2 Hz (5~100 s)中每一个频段的INS进行平均, 该频段可以避免由生理信号, 例如心跳(~1 Hz)、呼吸(0.2~0.3 Hz) (Kamran & Hong, 2014; Pierro, Hallacoglu, Sassaroli, Kainerstorfer, & Fantini, 2014)造成的脑间活动同步性。将平均以后的值进行Fisher Z转换。对每一个频段下22个通道的INS分别进行配对样本t检验, p < 0.005。结果显示, 惩罚条件下0.13~0.2 Hz、0.03~0.04 Hz的INS显著高于无惩罚条件, 但无惩罚条件下, 没有频段的INS高于惩罚条件(如图2所示)。因此, 选取0.13~0.2 Hz、0.03~0.04 Hz为研究频段。使用配对样本t检验分别考察这两个频段下每一个通道INS条件间的差异, 对p值进行FDR校正(Benjamini & Hochberg, 1995), 并生成t值热图, 通过xjview工具箱(
图2
3 结果
3.1 行为结果
图3
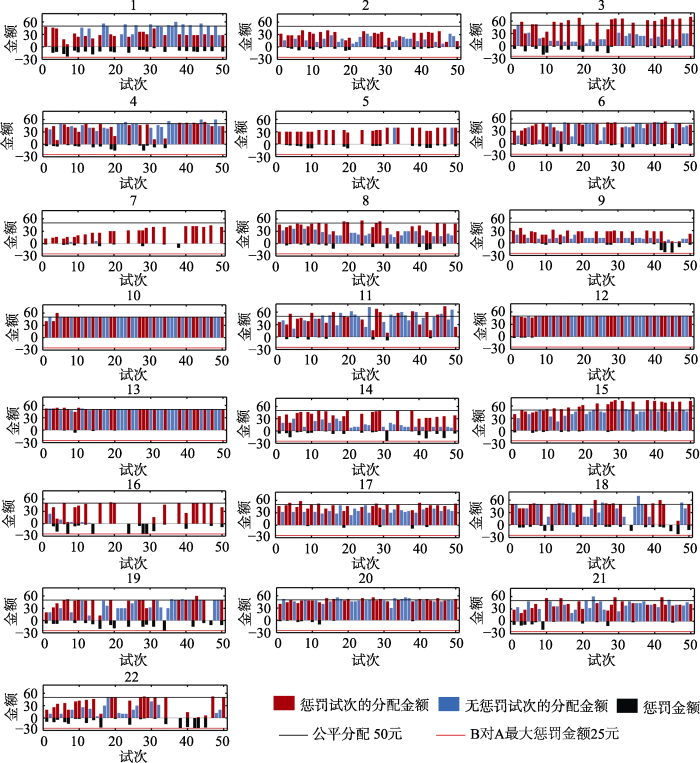
图4
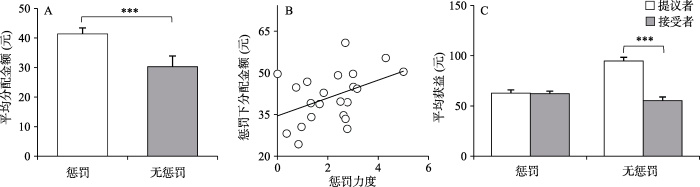
惩罚力度和惩罚下平均分配金额的相关分析的结果显示, 惩罚力度越大, 惩罚条件下提议者的分配金额越高, r = 0.43, p = 0.04 (如图4B所示)。另外, 对不同实验条件下提议者和接受者的平均获益进行配对样本t检验, 发现惩罚条件下, 提议者(62.76 ± 14.53)和接受者(62.22 ± 11.18)的平均获益没有显著差异, t(21) = 0.15, p = 0.88, Cohen's d = 0.03, 95%CI: -7~8.07; 而无惩罚条件下, 提议者的平均获益(94.72 ± 17)显著高于接受者(55.27 ± 17), t(21) = 5.44, p < 0.001, 95%CI: 24.38~54.53 (如图4C所示)。
3.2 fNIRS结果
分别考察0.13~0.2 Hz和0.03~0.04 Hz频段脑间活动同步性条件间的差异。结果显示, 当频段为0.13~0.2Hz时, 通道12和15惩罚条件下的INS显著高于无惩罚条件, 通道12: t(21) = 3.56, p = 0.04, Cohen's d = 0.76, 95%CI: 0.01~0.05; 通道15: t(21) = 3.45, p = 0.03, Cohen's d = 0.74, 95%CI: 0.01~0.05, p为FDR校正后(如图5A&B所示)。斯皮尔曼等级相关显示, 通道12的惩罚条件与无惩罚条件分配金额的差值与INS的差值呈正相关, ρ = 0.45, p = 0.04 (如图5C所示)。
图5
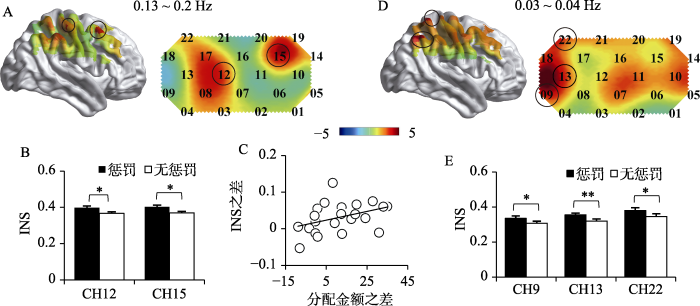
图5
通道12和15在0.13~0.2 Hz频段内INS条件间的差异(A&B); 惩罚与无惩罚条件下分配金额之差与通道12在0.13~0.2 Hz频段内的INS之差的相关性(C); 通道9、13和22在0.03~0.04 Hz频段内INS条件间的差异(D&E)
当频段为0.03~0.04 Hz时, 通道9、13和22惩罚条件下的INS显著高于无惩罚条件, 通道9: t(21) = 2.99, p = 0.05, Cohen's d = 0.64, 95%CI: 0.01~0.05; 通道13: t(21) = 4.21, p = 0.01, Cohen's d = 0.90, 95%CI: 0.02~0.05; 通道22: t(21) = 3.56, p = 0.02, Cohen's d = 0.76, 95%CI: 0.01~0.05, p为FDR校正后(如图5D&5E所示)。
4 讨论
本研究采用真实的人-人互动情境下的最后通牒博弈任务, 结合基于fNIRS的超扫描技术, 从群体互动视角下探究了人际公平形成的脑机制。行为结果显示, 惩罚下个体的分配金额更高, 且更接近平均分配。面对不公平分配, 接受者的惩罚力度越大, 提议者的分配越趋向平均分配。fNIRS的结果显示, 惩罚下提议者和接受者右侧背外侧前额叶皮层和“心理理论”相关脑区的脑间活动同步性(INS)增强, 且顶下小叶的脑间活动同步性的增强与分配金额的差异(惩罚-无惩罚)具有显著的相关关系。
研究中设置了两个实验条件, 惩罚条件和无惩罚条件。惩罚下, 接受者面对不公平分配时可以采取惩罚措施以纠正提议者的违规行为; 无惩罚下, 接受者没有惩罚权力, 只能接受提议者的任何分配方案。研究结果验证了惩罚对于促进提议者遵从公平规范的有效性和必要性。但与前人研究结果
(Spitzer et al., 2007)相比, 无惩罚下提议者的分配金额提高了20元。这可能是因为本研究中提议者和接受者在惩罚和无惩罚条件下进行了多轮分配, 导致两个实验条件相互有影响, 使得提议者的分配行为也受到了潜在的惩罚威胁的影响。我们还发现接受者面对不公平分配时的惩罚力度越大, 对促进提议者遵守公平规范的效应越明显。社会公平的形成一方面需要个体自觉遵从行为规范, 另一方面对违规行为也应做出合理的惩罚。如果接受者利益短视, 没有适度地对违规行为进行惩罚, 就会导致提议者为利益最大化, 保持不公分配, 甚至继续降低分配金额以试探接受者的底线。另外, 当不公分配已经发生时, 惩罚也是消除个体间差异的一种方式。研究结果显示, 惩罚下提议者和接受者最终的平均收益没有显著的差异。可能的原因有两点, 一是人们普遍厌恶不公平的分配, 其中既包括劣势不公平厌恶(自己比别人获得的少), 也包含优势不公平厌恶(自己比别人获得多) (Fehr & Schmidt, 1999; Gao et al., 2018)。二是适度的惩罚对于提议者已经具有威慑作用, 接受者也需尽可能减少惩罚金额以求收益最大化。
fNIRS的结果显示, 惩罚下提议者和接受者右侧背外侧前额叶皮层(DLPFC, 通道15)、右侧顶下小叶(IPL, 通道12和13)和右侧颞-顶联合区(TPJ, 通道9)的脑间活动同步性显著增强。以往研究显示, 右侧DLPFC在经济博弈中与认知控制、目标维持相关(Miller & Cohen, 2001; Sanfey, Rilling, Aronson, Nystrom, & Cohen, 2003)。在最后通牒博弈任务中, 提议者做出公平分配以及接受者拒绝不公平分配时, 右侧背外侧前额叶均会显著激活(Knoch, Gianotti, Baumgartner, & Fehr, 2010; Weiland, Hewig,Hecht, Mussel, & Miltner, 2012)。在惩罚条件下, 为避免因惩罚遭受损失, 提议者会趋向公平分配; 而接受者从长远利益出发, 会“自损八百”以促使提议者遵守公平规范。在这样的经济博弈中, 互动双方需要抑制优势反应以获得最大收益。由此惩罚下提议者和接受者右侧背外侧前额叶皮层神经活动的一致性也反映了这一共同的心理现象。
在最后通牒任务中, 提议者做出分配的时候往往会进行策略性的思考(strategic considerations), 对当前分配可能导致的接受者的情绪和行为反应进行估计(Weiland et al., 2012)。同样, 当接受者拥有惩罚权力来抑制不公平行为时, 也需对惩罚后提议者的行为反应加以推断。这种对他人心理状态进行解释和预测的认知活动称之为心理理论(mentalizing) (Corbetta, Patel, & Shulman, 2008; Gallagher & Frith, 2003)。本研究中, 我们发现惩罚下与“心理理论”相关脑区的INS显著增强, 包括右侧顶下小叶(IPL)和颞-顶联合区(TPJ)。IPL是镜像神经元系统的组成部分, 研究发现镜像神经元系统是社会认知重要的神经机制, 可以促进个体自我加工与社会理解(Casartelli & Molteni, 2014; Dapretto et al., 2006; Marsh & Hamilton, 2011)。右侧TPJ也是心理理论加工的重要脑区, 大量的研究证据表明右侧TPJ不仅与推断他人信念有关(Perner, Aichhorn, Kronbichler, Staffen, & Ladurner, 2006), 还涉及表征他人行为意图、整合意图与行为结果等(Koster-Hale, Saxe, Dungan, & Young, 2013; Young & Saxe, 2008)。“心理理论”相关的INS广泛存在于各种形式的社会互动中, 例如联合注意(Bilek et al., 2015)、同步唱歌(Osaka et al., 2015)、教学过程中的师生互动(Zheng et al., 2018)、人际交谈(Dai et al., 2018; Jiang et al., 2012)等, 且INS越强, 个体间的合作或互动越好。因此, INS可以作为衡量人际互动水平和共享意图的客观指标, 而惩罚下顶下小叶INS的增量与分配金额增量的正相关可能反应了惩罚下人际公平的提高伴随着博弈中互动双方更强的心理交互。
本研究首次尝试采用超扫描技术, 从社会互动视角下探究人际公平形成的神经机制, 并得出了一些结果, 但是研究仍存在不足之处。第一, 研究发现, 有些个体无论是惩罚条件还是无惩罚条件始终都能遵从公平规范; 而有些个体无惩罚条件下几乎不做任何分配, 这可能是个体之间自利水平的不同导致的。因此, 未来研究在探究人际公平问题的时候, 可以将人格特质纳入考察。第二, 遵从公平规范是一个复杂的社会认知过程, 不仅涉及大脑皮层脑区, 也与皮层下脑组织(例如脑岛、杏仁核等)紧密相关, 但是功能近红外光谱技术无法对皮层下的神经信号进行探测, 未来研究者们可以采纳高密度EEG源成像技术(Seeber et al., 2019)或者fMRI技术, 在保证研究生态效度的同时, 更全面地揭示人际公平形成的脑-脑机制。
5 结论
本研究首次结合修改版的最后通牒博弈任务和超扫描技术探讨了社会互动中人际公平形成的脑-脑机制, 本研究发现惩罚促进人际公平形成与互动双方前额叶、颞-顶联合区等部位更强的脑间活动同步有关。
参考文献
Inhibition and the right inferior frontal cortex
DOI:10.1016/j.tics.2004.02.010
Magsci
[本文引用: 1]

<h2 class="secHeading" id="section_abstract">Abstract</h2><p id="">It is controversial whether different cognitive functions can be mapped to discrete regions of the prefrontal cortex (PFC). The localisationist tradition has associated one cognitive function – inhibition – by turns with dorsolateral prefrontal cortex (DLPFC), inferior frontal cortex (IFC), or orbital frontal cortex (OFC). Inhibition is postulated to be a mechanism by which PFC exerts its effects on subcortical and posterior-cortical regions to implement executive control. We review evidence concerning inhibition of responses and task-sets. Whereas neuroimaging implicates diverse PFC foci, advances in human lesion-mapping support the functional localization of such inhibition to right IFC alone. Future research should investigate the generality of this proposed inhibitory function to other task domains, and its interaction within a wider network.</p>
Dorsolateral and ventromedial prefrontal cortex orchestrate normative choice
Controlling the false discovery rate: A practical and powerful approach to multiple testing
Information flow between interacting human brains: Identification, validation, and relationship to social expertise
Where there is a goal, there is a way: What, why and how the parieto-frontal mirror network can mediate imitative behaviours
The reorienting system of the human brain: From environment to theory of mind
DOI:10.1016/j.neuron.2008.04.017
Magsci
[本文引用: 1]
<p id="">Survival can depend on the ability to change a current course of action to respond to potentially advantageous or threatening stimuli. This “reorienting” response involves the coordinated action of a right hemisphere dominant ventral frontoparietal network that interrupts and resets ongoing activity and a dorsal frontoparietal network specialized for selecting and linking stimuli and responses. At rest, each network is distinct and internally correlated, but when attention is focused, the ventral network is suppressed to prevent reorienting to distracting events. These different patterns of recruitment may reflect inputs to the ventral attention network from the locus coeruleus/norepinephrine system. While originally conceptualized as a system for redirecting attention from one object to another, recent evidence suggests a more general role in switching between networks, which may explain recent evidence of its involvement in functions such as social cognition.</p>
NIRS-based hyperscanning reveals increased interpersonal coherence in superior frontal cortex during cooperation
DOI:10.1016/j.neuroimage.2011.09.003
Magsci
[本文引用: 2]
We used Near-Infrared Spectroscopy (NIRS) to simultaneously measure brain activity in two people while they played a computer-based cooperation game side by side. Inter-brain activity coherence was calculated between the two participants. We found that the coherence between signals generated by participants' right superior frontal cortices increased during cooperation, but not during competition. Increased coherence was also associated with better cooperation performance. To our knowledge, this work represents the first use of a single NIRS instrument for simultaneous measurements of brain activity in two people. This study demonstrates the use of NIRS-based hyperscanning in studies of social interaction in a naturalistic environment. (C) 2011 Elsevier Inc. All rights reserved.
Neural mechanisms for selectively tuning in to the target speaker in a naturalistic noisy situation
Understanding emotions in others: Mirror neuron dysfunction in children with autism spectrum disorders
Dorsal striatum responses to reward and punishment: Effects of valence and magnitude manipulations
Cooperation and punishment in public goods experiments
A theory of fairness, competition, and cooperation
Neural signatures of fairness-related normative decision making in the ultimatum game: A coordinate-based meta-analysis
The Ultimatum Game and the brain: A meta-analysis of neuroimaging studies
Functional imaging of ‘theory of mind’
Distinguishing neural correlates of context-dependent advantageous-and disadvantageous- inequity aversion
Application of the cross wavelet transform and wavelet coherence to geophysical time series
An experimental analysis of ultimatum bargaining
Brain-to-brain coupling: A mechanism for creating and sharing a social world
DOI:10.1016/j.tics.2011.12.007
Magsci
[本文引用: 1]
Cognition materializes in an interpersonal space. The emergence of complex behaviors requires the coordination of actions among individuals according to a shared set of rules. Despite the central role of other individuals in shaping one's mind, most cognitive studies focus on processes that occur within a single individual. We call for a shift from a single-brain to a multi-brain frame of reference. We argue that in many cases the neural processes in one brain are coupled to the neural processes in another brain via the transmission of a signal through the environment. Brain-to-brain coupling constrains and shapes the actions of each individual in a social network, leading to complex joint behaviors that could not have emerged in isolation.
Costly punishment across human societies
Between-brain connectivity during imitation measured by fNIRS
DOI:10.1016/j.neuroimage.2012.06.028
Magsci
[本文引用: 1]
The present study aimed to step into two-person neuroscience by investigating the hemodynamic correlates of between-brain connectivity involved in imitation and its dependency on pacing stimuli. To test this approach, we used wireless functional near-infrared spectroscopy (fNIRS) to record simultaneously during imitation performance of a paced finger-tapping task (PFT) in two subjects over premotor cortices (PMC). During the imitation (IM) condition, a model and an imitator were recorded while tapping in synchrony with auditory stimuli separated by a constant interval (stimulus-paced mode, St-P), followed by tapping without the pacing stimulus (self-paced mode, Se-P). During the control (CO) condition, each subject (single 1 and 2) performed the PFT task with the same pacing mode pattern, but alone without reference to each other.<br/>Using wavelet transform coherence (WTC) analysis evaluating functional connectivity between brains, we found (1) that IM revealed a larger coherence increase between the model and the imitator as compared to the CO condition. (2) Within the IM condition, a larger coherence increase was found during Se-P as compared to St-P mode. Using Granger-causality (G-causality) analysis evaluating effective connectivity between brains, we found (3) that IM revealed larger G-causality as compared to the CO condition and (4) that within the IM condition, the signal of the model G-caused that of the imitator to a greater extent as compared to vice versa.<br/>Our findings designate fNIRS as suitable tool for monitoring between-brain connectivity during dynamic interactions between two subjects and that those measurements might thereby provide insight into activation patterns not detectable using typical single-person experiments. Overall, the results of the present study demonstrate the potential of simultaneously assessing brain hemodynamics in interacting subjects in several research areas where social interactions are involved. (C) 2012 Elsevier Inc. All rights reserved.
Functional near-infrared optical imaging: Utility and limitations in human brain mapping
Neural synchronization during face-to-face communication
DOI:10.1523/JNEUROSCI.2926-12.2012
Magsci
[本文引用: 1]
Although the human brain may have evolutionarily adapted to face-to-face communication, other modes of communication, e. g., telephone and e-mail, increasingly dominate our modern daily life. This study examined the neural difference between face-to-face communication and other types of communication by simultaneously measuring two brains using a hyperscanning approach. The results showed a significant increase in the neural synchronization in the left inferior frontal cortex during a face-to-face dialog between partners but none during a back-to-back dialog, a face-to-face monologue, or a back-to-back monologue. Moreover, the neural synchronization between partners during the face-to-face dialog resulted primarily from the direct interactions between the partners, including multi-modal sensory information integration and turn-taking behavior. The communicating behavior during the face-to-face dialog could be predicted accurately based on the neural synchronization level. These results suggest that face-to-face communication, particularly dialog, has special neural features that other types of communication do not have and that the neural synchronization between partners may underlie successful face-to-face communication.
Reduction of physiological effects in fNIRS waveforms for efficient brain-state decoding
A neural marker of costly punishment behavior
Studying the neurobiology of social interaction with transcranial direct current stimulation—the example of punishing unfairness
Diminishing reciprocal fairness by disrupting the right prefrontal cortex
Decoding moral judgments from neural representations of intentions
Role of the right inferior frontal gyrus in turn-based cooperation and competition: A near-infrared spectroscopy study
A neural mechanism of strategic social choice under sanction-induced norm compliance
Dissociation of mirroring and mentalising systems in autism
DOI:10.1016/j.neuroimage.2011.02.003
Magsci
[本文引用: 1]
<h2 class="secHeading" id="section_abstract">Abstract</h2><p id="sp0030">The role of mirror neuron systems and mentalising systems in causing poor social and communication skills in individuals with autistic spectrum conditions is hotly debated. We studied 18 adults with autistic spectrum conditions in comparison to 19 age and IQ matched typical individuals. Behavioural assessments revealed difficulties in mental state attribution and action comprehension in the autism sample. We examined brain responses when observing rational and irrational hand actions, because these actions engage mirror and mentalising components of the social brain respectively.</p><p id="sp0035">Both typical and autistic participants activated the left anterior intraparietal sulcus component of the mirror system when viewing hand actions compared to moving shapes. The typical but not autistic participants activated the posterior mid cingulate cortex/supplementary motor area and bilateral fusiform cortex when viewing hand actions. When viewing irrational hand actions, the medial prefrontal cortex of typical participants deactivated but this region did not distinguish the different stimuli in autistic participants. These results suggest that parietal mirror regions function normally in autism, while differences in action understanding could be due to abnormal function of cingulate, fusiform and medial prefrontal regions. Thus, brain regions associated with mirroring and mentalising functions are differentially affected in autistic spectrum conditions.</p><h4 id="secGabs_N37b60e90N3778f7d0">Research highlights</h4><p>? Adults with/without autism observed rational and irrational actions during fMRI. ? Anterior intraparietal sulcus showed equivalent responses in both groups. ? Supplementary motor area responded to action observation only in typicals. ? Medial prefrontal cortex differentiated action rationality only in typicals. ? Mirroring (aIPS) and mentalising (mPFC) brain area dissociate in autism.</p>
An integrative theory of prefrontal cortex function
Hyperscanning: Simultaneous fMRI during linked social interactions
Interpersonal frontopolar neural synchronization in group communication: An exploration toward fNIRS hyperscanning of natural interactions
Abstract reward and punishment representations in the human orbitofrontal cortex
How two brains make one synchronized mind in the inferior frontal cortex: fNIRS-based hyperscanning during cooperative singing
Cooperation in lovers: An fNIRS-based hyperscanning study
Interpersonal synchronization of inferior frontal cortices tracks social interactive learning of a song
Thinking of mental and other representations: The roles of left and right temporo-parietal junction
Validation of a novel hemodynamic model for coherent hemodynamics spectroscopy (CHS) and functional brain studies with fNIRS and fMRI
Brain-to-brain synchrony in parent-child dyads and the relationship with emotion regulation revealed by fNIRS-based hyperscanning
Changing social norm compliance with noninvasive brain stimulation
DOI:10.1126/science.1241399
Magsci
[本文引用: 1]
All known human societies have maintained social order by enforcing compliance with social norms. The biological mechanisms underlying norm compliance are, however, hardly understood. We show that the right lateral prefrontal cortex (rLPFC) is involved in both voluntary and sanction-induced norm compliance. Both types of compliance could be changed by varying the neural excitability of this brain region with transcranial direct current stimulation, but they were affected in opposite ways, suggesting that the stimulated region plays a fundamentally different role in voluntary and sanction-based compliance. Brain stimulation had a particularly strong effect on compliance in the context of socially constituted sanctions, whereas it left beliefs about what the norm prescribes and about subjectively expected sanctions unaffected. Our findings suggest that rLPFC activity is a key biological prerequisite for an evolutionarily and socially important aspect of human behavior.
The neural basis of economic decision-making in the ultimatum game
A new methodical approach in neuroscience: Assessing inter- personal brain coupling using functional near-infrared imaging (fNIRI) hyperscanning
Subcortical electrophysiological activity is detectable with high-density EEG source imaging
A dual-fMRI investigation of the iterated Ultimatum Game reveals that reciprocal behaviour is associated with neural alignment
The neural signature of social norm compliance
DOI:10.1016/j.neuron.2007.09.020
Magsci
[本文引用: 5]
<p id="">Compliance with social norms requires neural signals related both to the norm and to deviations from it. Recent work using economic games between two interacting subjects has uncovered brain responses related to norm compliance and to an individual's strategic outlook during the exchange. These brain responses possess a provocative relationship to those associated with negative emotional outcomes, and hint at computational depictions of emotion processing.</p>
Interpersonal brain synchronization in the right temporo-parietal junction during face-to-face economic exchange
Neural correlates of fair behavior in interpersonal bargaining
Neural basis of increased costly norm enforcement under adversity
DOI:10.1093/scan/nst187
Magsci
[本文引用: 1]
Humans are willing to punish norm violations even at a substantial personal cost. Using fMRI and a variant of the ultimatum game and functional magnetic resonance imaging, we investigated how the brain differentially responds to fairness in loss and gain domains. Participants (responders) received offers from anonymous partners indicating a division of an amount of monetary gain or loss. If they accept, both get their shares according to the division; if they reject, both get nothing or lose the entire stake. We used a computational model to derive perceived fairness of offers and participant-specific inequity aversion. Behaviorally, participants were more likely to reject unfair offers in the loss (vs gain) domain. Neurally, the positive correlation between fairness and activation in ventral striatum was reduced, whereas the negative correlations between fairness and activations in dorsolateral prefrontal cortex were enhanced in the loss domain. Moreover, rejection-related dorsal striatum activation was higher in the loss domain. Furthermore, the gain-loss domain modulates costly punishment only when unfair behavior was directed toward the participants and not when it was directed toward others. These findings provide neural and computational accounts of increased costly norm enforcement under adversity and advanced our understanding of the context-dependent nature of fairness preference.
The context-dependency of fairness processing: Evidence from ERP study
DOI:
Magsci
In human society, sharing losses is at least as common as sharing gains. Although the psychological and neural processes underlying the latter have been investigated in depth, those related to the former are not clear. Our recent study demonstrates an increased demand for fairness under adversity (e.g. loss sharing). Here we investigated how our brain encodes unfairness in the loss and gain domains using event-related potentials (ERP) technique. We adopted the Ultimatum Game (UG) to probe the processes related to fairness consideration in either gain or loss domain. In UG, two players, the proposer and the responder, bargain on how to divide a certain amount of money endowed by the experimenter. The proposer suggests a division policy, on which the responder evaluates and decides whether to accept. Upon acceptance, the money is divided as suggested; while rejection results in both players going empty-handed. Participants, as responders, were required to decide whether to accept an offer that was either fair (equal or nearly equal division) or unfair in both gain and loss domain. Offers were either made by the human partner or by the computer partner. Behavioral results replicated our previous findings that the rejection rate of unfair offers was higher in the loss than in the gain domain. ERP results revealed that the N1 amplitude was more pronounced for human partners compared with computer partners, however, this effect was only observed in the gain domain. When interacting with computer partners, unfair offers and offers in the loss domain were associated with larger N350 compared with fair offers and offers in the gain domain, and offers in the gain domain elicited larger P2 than offers in the loss domain, whereas fair offers and offers in the gain domain were associated with larger LPP than unfair offers and offers in the loss domain. In addition, these differences in ERP responses were diminished when the interacting partners were humans. These findings suggest that fairness processing is modulated by the property of the partner and gain-loss domain. In human-computer interaction, unfair offers and offers in the loss domain elicit more inhibition and conflict resolving process, while fair offers and offers in the gain domain are more motivationally significant to human. The present findings support the view that fairness processing is context-dependent, in which factors like gain-loss domain and the property of the partner play a role.
公平加工的情境依赖性: 来自 ERP 的证据
公平是人类社会生活的重要概念。大量研究采用最后通牒博弈, 发现人们具有不公平厌恶倾向, 即宁愿牺牲个人的经济利益, 也要拒绝不公平的分配提议。已有研究表明, 损失情境会增强不公平厌恶, 但其神经机制尚不清楚。本实验采用ERP技术, 运用最后通牒博弈范式, 考察两个情境因素:域(损失或获益)和博弈对象(人或计算机)对公平加工的影响。发现损失域下对不公平分配的拒绝率更高, 而博弈对象对决策行为无影响; 获益域下, 对家为人比对家为计算机诱发了更负的N1; 人机博弈时, 获益域比损失域诱发了更大的P2和LPP, 损失域比获益域、不公平提议比公平提议诱发了更大的N350, 而公平提议比不公平提议诱发了更大的LPP; 人际博弈时, 这些差异均不显著。这些结果表明, 对分配提议的大脑加工受博弈对象的调节, 人际博弈时, 对损益域、公平与不公平提议的加工类似, 而人机博弈时, 损失域和不公平提议涉及更多的抑制加工和冲突解决, 获益域和公平提议则更富动机性意义, 证实公平加工具有情境依赖性。
BrainNet Viewer: A network visualization tool for human brain connectomics
NIRS-SPM: Statistical parametric mapping for near-infrared spectroscopy
DOI:10.1016/j.neuroimage.2008.08.036
Magsci
[本文引用: 1]
<h2 class="secHeading" id="section_abstract">Abstract</h2><p id="">Near infrared spectroscopy (NIRS) is a non-invasive method to measure brain activity via changes in the degree of hemoglobin oxygenation through the intact skull. As optically measured hemoglobin signals strongly correlate with BOLD signals, simultaneous measurement using NIRS and fMRI promises a significant mutual enhancement of temporal and spatial resolutions. Although there exists a powerful statistical parametric mapping tool in fMRI, current public domain statistical tools for NIRS have several limitations related to the quantitative analysis of simultaneous recording studies with fMRI. In this paper, a new public domain statistical toolbox known as NIRS-SPM is described. It enables the quantitative analysis of NIRS signal. More specifically, NIRS data are statistically analyzed based on the general linear model (GLM) and Sun's tube formula. The <em>p</em>-values are calculated as the excursion probability of an <em>inhomogeneous</em> random field on a representation manifold that is dependent on the structure of the error covariance matrix and the interpolating kernels. NIRS-SPM not only enables the calculation of activation maps of oxy-, deoxy-hemoglobin and total hemoglobin, but also allows for the super-resolution localization, which is not possible using conventional analysis tools. Extensive experimental results using finger tapping and memory tasks confirm the viability of the proposed method.</p>
The neural basis of belief encoding and integration in moral judgment
DOI:10.1016/j.neuroimage.2008.01.057
Magsci
[本文引用: 1]
<h2 class="secHeading" id="section_abstract">Abstract</h2><p id="">Moral judgment in the mature state depends on “theory of mind”, or the capacity to attribute mental states (e.g., beliefs, desires, and intentions) to moral agents. The current study uses functional magnetic resonance imaging (fMRI) to investigate the cognitive processes for belief attribution in moral judgment. Participants read vignettes in a 2 × 2 × 2 design: protagonists produced either a negative or neutral outcome, based on the belief that they were causing the negative outcome or the neutral outcome; presentation of belief information either preceded or followed outcome information. In each case, participants judged the moral permissibility of the action. The results indicate that while the medial prefrontal cortex is recruited for processing belief valence, the temporo-parietal junction and precuneus are recruited for processing beliefs in moral judgment via two distinct component processes: (1) encoding beliefs and (2) integrating beliefs with other relevant features of the action (e.g., the outcome) for moral judgment.</p>
Enhancement of teaching outcome through neural prediction of the students' knowledge state

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}