

1 前言
伴随强烈恐惧记忆往往是临床上许多精神障碍疾病的主要症状表现, 如创伤后应激障碍(PTSD, post- traumatic stress disorder)、焦虑症(anxiety disorder)、恐惧症(phobia)等。目前临床心理治疗方法存在的重要问题之一是不能有效的抑制恐惧记忆的复发, 恐惧症状复发比率高 (Choy, Fyer, & Lipsitz, 2007)。因此寻求更加有效擦除不良恐惧记忆并且抑制恐惧记忆复发的方法, 一直是国内外研究者的研究焦点与主要目标。
当前实验室研究恐惧主要采用基于巴甫洛夫条件反射的条件性恐惧模型, 将条件刺激(conditioned stimulus, CS, 中性刺激, 如图片、声音、光)与非条件刺激(unconditioned stimulus, US, 厌恶刺激, 如电击)配对多次强化形成(CS-US)连接, 使得单独出现条件刺激时会引发恐惧反应, 称为恐惧习得。之后如果条件刺激多次出现始终没有配对非条件刺激(CS-no US)时, 条件刺激引发的恐惧反应会逐渐减少甚至消失, 这就是传统的恐惧消退模式(Davis, Walker, & Myers, 2003)。传统恐惧消退模式(如暴露疗法)一直是临床对PTSD等情绪障碍的应对策略, 但在许多情况下都会被重新诱发, 表现为自发恢复(Spontaneous recovery)、重建(Reinstatement)、续新(Renewal)和再习得(Reacquisition)等恐惧返回的行为特征(Bouton & Swartzentruber, 1991; Myers & Davis, 2007)。近年来研究者把焦点转向记忆的再巩固过程, 尝试用各种基于记忆再巩固的干预模式来消退恐惧记忆。
传统的记忆固化理论(perseveration consolidation theory), 认为记忆的形成是一次性的过程, 经过编码、固化和稳定阶段, 一旦巩固下来就难以改变(McGaugh, 2000)。而记忆再巩固理论(reconsolidation theory)认为, 记忆的形成不是一蹴而就的, 是一个不断提取并再次储存的过程。记忆每次被提取激活后会进入不稳定的状态, 要经过新一轮的巩固过程记忆才能保持, 期间需要有蛋白质的合成。如果在这个阶段进行药物或行为干预, 会使原有的记忆受到干扰从而可能改写(rewrite)或清除(erase)原有的记忆(Lee, Everitt, & Thomas, 2004; Lee, Nader, & Schiller, 2017; Nader, Schafe, & LeDoux, 2000)。基于再巩固理论, Monfils, Cowansage, Klann和LeDoux (2009)使用动物条件恐惧模型, 首次提出一种安全、无伤害无入侵性的方法来消除恐惧记忆, 即提取消退(retrieval-extinction)范式。该范式通过单独呈现的CS条件刺激激活原始记忆使其进入不稳定状态, 在这个再巩固时间窗内进行传统消退将安全信息融入到原始记忆中从而减弱或抑制恐惧复发(Monfils et al., 2009)。Schiller等(2010)成功地将这一范式应用于人类被试, 证明了在不使用药物的情况下使用行为训练能够破坏人类恐惧记忆, 有效阻止恐惧复发, 且效果持续一年以上(Schiller et al., 2010)。提取消退范式不仅对条件性恐惧记忆消除有效, 对成瘾记忆这一非常强烈的记忆也具有擦除作用, 我国研究者首次将此范式应用于药物成瘾消退研究上(Xue et al., 2012), 证明了该模式可以有效消退吸毒成瘾个体的药物依赖并减少复吸。之后又针对线索特异性问题, 进一步开发出非条件性刺激(US)提取消退模式(Liu et al., 2014; Luo et al., 2015)。但是目前关于提取消退范式的研究还存在争议, 有些研究出现了不一致的结果(Ishii et al., 2015; Kindt & Soeter, 2013; Soeter & Kindt, 2011)。有研究者认为可能是因为这类破坏记忆再巩固的干预范式作用存在一个前提条件, 只有满足一定的条件记忆才会进入再巩固, 这类条件称为再巩固的边界条件(boundary conditions)。大量研究已经证实了一系列边界条件的存在, 如记忆年龄、记忆强度、提取强度等(Auber, Tedesco, Jones, Monfils, & Chiamulera, 2013; Nader, 2015; Schwabe, Nader, & Pruessner, 2014)。
最近有研究者提出预期错误(Prediction Error, PE)也是记忆再巩固的边界条件(Fernández, Boccia, & Pedreira, 2016; Li & McNally, 2014)。预期错误最早由Rescorla和Wagner (1972)研究巴甫洛夫条件反射中强化与非强化效果变化时提出, 是指先前认知的信息与现在认知的信息之间出现差异和不匹配, 研究发现预期错误能够影响条件反射的连结效果(Rescorla & Wagner, 1972)。预期错误又分负性预期错误(Negative PE)和正性预期错误(Positive PE)两种类型, 负性PE是指预期会出现结果没有, 正性PE是指预期不会出现结果有(Beckers & Kindt, 2017)。目前学界普遍认为, 记忆再巩固过程实际上是通过允许新信息整合进原先建立的记忆痕迹(memory trace)中, 目的是保持记忆的相关性(Exton-McGuinness, Lee, & Reichelt, 2015)。当提取时没有偏离原先记忆的信息出现时, 就不需要更新记忆, 因为这种情况下没有必要冒着记忆被干扰的风险进行再巩固。这也符合记忆自适应原则, 现实生活中我们也并不是每次回忆事件时被提取的记忆就会变得不稳定进入再巩固。所以只有当记忆提取时出现某些违反预期的新信息时记忆才会变得不稳定, 再巩固才有必要开启来实现记忆自适应。Sevenster, Beckers和Kindt (2013)的研究将预期错误用于记忆再巩固和恐惧记忆消退方面, 发现可以用预期错误来引发记忆再巩固。在该研究中, 预期错误被定义为记忆提取阶段与习得阶段内容不匹配, 在提取阶段操纵预期错误, 进而利用β-肾上腺素能受体拮抗剂(propranolol)验证记忆是否进入再巩固, 结果发现只有出现预期错误, 恐惧记忆才会经历再巩固过程(Sevenster et al., 2013)。越来越多的证据表明, 预期错误在引发记忆再巩固过程中起到非常关键的作用, 但是对于预期错误是不是记忆进入再巩固的必要条件和开启因素还需进行进一步的研究。
在以往关于预期错误的研究中, 研究者们大多数采用药物等侵入性干预模式来验证记忆是否经历再巩固, 较少研究将预期错误应用于非侵入性的提取消退范式中。而且现有研究很少去探讨预期错误作为边界条件对记忆再巩固的作用, 也很少有研究试图整合预期错误的类型与数量大小去探究多大程度的预期错误开启再巩固效果最好。有研究者认为预期错误过大可能不会使记忆进入再巩固, 因为现有信息与原先记忆很不匹配会被认为是与原先记忆无关的全新信息(Díaz-Mataix, Ruiz Martinez, Schafe, LeDoux, & Doyère, 2013; Sevenster, Beckers, & Kindt, 2014)。而且在现实情境中, 预测危险的信号可能是有多个不同的线索, 也可能来自多个不同的感观通道, 将条件刺激设置成多感官复合模型(声音 + 图片)能够更加贴近现实情境(庄楚群等, 2017)。因此, 本研究将尝试把预期错误应用到复合记忆模型的提取消退范式中, 采用多感官复合模型(声音+图片)作为条件刺激形成较为复杂的条件性恐惧记忆, 在提取阶段通过改变CS-US配对规则来操纵预期错误的类型以及数量, 探讨预期错误是不是启动记忆再巩固过程的必要条件, 以及预期错误的大小是否会影响记忆进入再巩固过程。进一步验证和丰富提取消退这一恐惧记忆消退模式, 提高提取消退范式在心理治疗中的临床应用价值。
2 方法
2.1 被试
被试为86名在校大学生, 通过自愿报名的方式参加实验, 有一定的实验报酬。被试均为右利手, 无躯体疾病及精神障碍病史, 视力或矫正视力正常, 无色盲色弱, 听力正常, 最近没有鼻塞或咳嗽症状, 且之前没有参加类似的实验。整个实验过程与被试招募标准均通过科研伦理委员会审核。实验前一天通知被试保持良好睡眠, 实验三天期间保持正常作息, 以排除实验被试偏差。实验前告知被试实验中会在手臂处接受轻微电击, 电击强度是经过科学评定的, 不会对人体造成任何伤害。实验中如果有任何不适症状可以随时自由退出,所有与被试有关的信息和数据都会被严格保密。被试了解情况后需在知情同意书上签字, 随后填写《状态-特质焦虑量表》。
被试随机分配进入4个组, 分别为“无预期错误提取消退组” (无PE组)、“单个负性预期错误提取消退组” (负性PE组)、“单个正性预期错误提取消退组” (正性PE组)和“多重预期错误提取消退组” (多重PE组)。实验过程中有2个被试由于在第一天无法习得刺激伴随电击的规律没有进行接下来的实验, 在数据分析中予以剔除, 故有效被试84人, 其中无PE组21人(5名男生), 负性PE组21人(5名男生), 正性PE组21人(4名男生), 多重PE组21人(2名男生)。被试年龄范围为18~24岁(M = 20, SD = 1)。状态-特质焦虑量表结果显示:4组被试在状态焦虑分量表(F(3,80) = 1.63, p > 0.05)和特质焦虑分量表(F(3,80) = 1.71, p > 0.05)上的得分均无显著差异。
2.2 实验材料
本实验条件刺激采用双线索图片和声音组成的复合刺激, 图片为两类不同颜色的立体几何图形组合, 图片亮度相同, 背景为白色。声音选取自经过大范围评定的本土化情绪声音库(刘涛生, 罗跃嘉, 马慧, 黄宇夏, 2006), 分别为铃声(愉悦度 = 5 ± 1.11, 唤醒度 = 5.92 ± 1.52, 优势度 = 5.08 ± 1.81)和钟声(愉悦度 = 5.08 ± 1.43, 唤醒度 = 4.58 ± 1.91, 优势度 = 4.68 ± 1.58), 均为中性声音。两种声音的响度相同(材料评定阶段, 有96.5%的人能清晰分辨这两种声音)。这两类图片与声音两两组合形成两类复合刺激。实验中复合刺激刺激呈现时间为8000 ms, 其中一种复合刺激呈现后不定时会伴随非条件刺激(电击), 作为CS+, 另一种条件刺激呈现后不会伴随非条件刺激, 作为CS-。非条件刺激采用电击仪电击被试右手腕部造成恐惧反应, 电击强度事先根据每个被试的承受能力进行评定, 均控制在每个被试可承受范围内且不会对人体造成伤害。每次电击持续时间为200 ms。为避免实验材料对被试间造成的影响, 实验中所有的材料都会进行项目平衡。
2.3 测量指标
使用Biopac 16通道生理记录仪(型号MP150)记录被试的皮肤电(skin conductance responses, SCR)信号(Lonsdorf et al., 2017)。实验中Ag/AgCl电极分别固定在被试左手无名指与食指末端, 电极另一端口连接生理记录仪的EDA100C模块上, 采样率为500 Hz。分析软件采用Biopac 16通道生理记录仪自带的AcqKnowledge 4.2软件, 处理方法参考Schiller团队采用的计算方法, 具体为:取CS呈现后500~7500 ms时间窗内的最高值和最低值, 两者之差为CS诱发的皮电值, 随后对所有被试的皮电值要进行范围校正, 分别除以每个被试的US诱发皮电值的平均值, 最后所有数据平方根化以减少分布的偏度(Schiller et al., 2010)。
2.4 实验程序
电击强度评定阶段:在第一天正式实验开始之前每个被试都需要接受电击强度评定以确保这个强度能够让被试感到很不舒服但能够忍受。电击强度的调节范围为10~50 V, 被试每次接受电击后需对电击造成的感受进行0~10评级(其中0为舒服、8为极度不舒服但能忍受、9为痛到不能忍受)。选取被试评定为等级8的电击强度作为该被试整个实验过程的电击强度。
实验材料采用E-prime 2.0软件编程呈现在同一电脑屏幕上(声音刺激通过防噪耳机连接佩戴于被试耳部)。每一试次首先在屏幕上呈现红色注视点“+”2000 ms提醒被试注意, 随后呈现条件刺激CS (声音+图片), 持续8000 ms。若CS需伴随US则电击会在CS消失前200 ms实施电击, 电击持续时间为200 ms, 随后一同消失。试次间隔(ITI)为8~10 s, 间隔期间屏幕呈现“请放松”字样, 确保被试的皮电值能降到标准水平(如图1所示)。正式实验过程分为3天, 第一天为恐惧习得阶段, 第二天为提取-消退阶段, 第三天为记忆复发测试阶段, 正式实验全程被试左手连接生理记录仪收集皮肤电, 右手连接电击仪。
图1
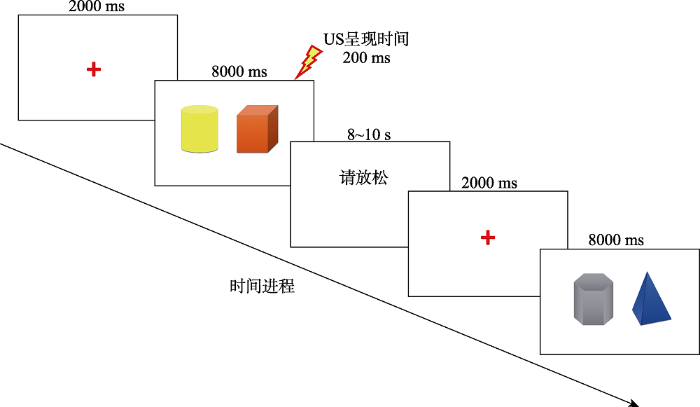
第一天恐惧习得阶段:采用电击次序建立学习规则(Sevenster et al., 2013)。CS+和CS-各呈现6次, 连续两次交替呈现, 即先呈现连续两个CS+, 再连续呈现两个CS-, 以此类推。其中无PE组与负性PE组中连续呈现的两个CS+中总是第一个不伴随电击, 第二个伴随电击。正性PE组与多重PE组中连续呈现的两个CS+中总是第一个伴随电击, 第二个不伴随电击。4个组CS-均不伴随电击, CS+的电击比率为50%, 各组的设置情况如图2所示(该图仅呈现CS+的出现次序)。该阶段要求被试全程集中注意力在电脑屏幕上, 并观察出刺激与电击之间的规律。
图2

第二天提取-消退阶段:参考Kindt等人设置预期错误的方法(Sevenster et al., 2013, 2014), 无PE组提取时单独呈现一个不伴随电击的CS+、负性PE组提取时连续呈现两个不伴随电击的CS+、正性PE组提取时连续呈现两个伴随电击的CS+、多重PE组提取时连续呈现4个不伴随电击的CS+。提取之后被试休息10 min, 随后进入消退阶段, 消退阶段随机呈现10个CS+和10个CS-, 均不伴随电击。该阶段要求被试全程集中注意力在电脑屏幕上, 不需要做任何反应。
第三天恐惧复发测试阶段:首先进行恐惧记忆自发恢复测试, 使用一个完整的消退序列进行消退,随机呈现8个CS+和8个CS-, 均不伴随电击, 以第二天最后一个试次的CS+与第三天第一个试次的CS+变化作为衡量恐惧自发恢复的指标。再消退结束后休息1 min, 随后进行恐惧记忆重建测试, 向被试连续呈现4个无预警的US电击, 每次电击持续200 ms, 间隔1000 ms。被试休息5 min后进行再次消退, 随机呈现7个CS+和7个CS-, 均不伴随电击, 以自发恢复测试的消退序列的最后一个CS+与重建测试的消退训练的第一个CS+的变化作为衡量恐惧重建的指标。具体设计见表1。该阶段要求被试全程集中注意力在电脑屏幕上, 不需要做任何反应。
表1 实验设计
| 第一天 | 第二天 | 第三天 | ||||||
|---|---|---|---|---|---|---|---|---|
| 习得3 CS+US | 无PE提取 | 10 min | 消退训练 | 自发恢复测试 | 1 min | 4US | 5 min | 重建测试 |
| 3 CS+ | 10 CS+消退 | 8 CS+ | 7 CS+ | |||||
| 6 CS- | 10 CS-消退 | 8 CS- | 7 CS- | |||||
| 习得 3 CS+US | 单个负性PE提取 | 10 min | 消退训练 | 自发恢复测试 | 1 min | 4US | 5 min | 重建测试 |
| 3 CS+ | 10 CS+消退 | 8 CS+ | 7 CS+ | |||||
| 6 CS- | 10 CS-消退 | 8 CS- | 7 CS- | |||||
| 习得 3 CS+US | 单个正性PE提取 | 10 min | 消退训练 | 自发恢复测试 | 1 min | 4US | 5 min | 重建测试 |
| 3 CS+ | 10 CS+消退 | 8 CS+ | 7 CS+ | |||||
| 6 CS- | 10 CS-消退 | 8 CS- | 7 CS- | |||||
| 习得 3 CS+US | 多重PE提取 | 10 min | 消退训练 | 自发恢复测试 | 1 min | 4US | 5 min | 重建测试 |
| 3 CS+ | 10 CS+消退 | 8 CS+ | 7 CS+ | |||||
| 6 CS- | 10 CS-消退 | 8 CS- | 7 CS- |
3 实验结果
对被试的皮肤电(SCR)数据, 以刺激类型(CS+/ CS-)和试次为被试内因素, 组别(无PE组、负性PE组、正性PE组和多重PE组)为被试间因素进行重复测量方差分析。
条件恐惧习得:重复测量方差分析结果显示, 刺激类型的主效应显著, F(1,80) = 58.15, p < 0.001, 偏η2 = 0.42; 试次的主效应显著, F(5,76) = 9.31, p < 0.001,偏η2 = 0.38; 刺激类型和试次的交互作用显著, F(5,76) = 2.88, p < 0.05, 偏η2 = 0.16, 说明CS+和CS-的关系从习得前期到习得后期发生显著变化。组别效应不显著, F(3,80) = 0.46, p = 0.71; 刺激类型和组别的交互作用也不显著, F(3,80) = 0.85, p = 0.43, 说明4组被试对CS+和CS-的恐惧反应不存在显著差异。再分别对每组后半部分(习得的最后3个试次SCR平均值)的CS+与CS-进行配对样本t检验, 结果显示, 无PE组差异显著, t(20) = 3.08, p < 0.05, d = 0.67; 负性PE组差异显著, t(20) = 2.57, p < 0.05, d = 0.56; 正性PE组差异显著, t(20) = 4.53, p < 0.001, d = 0.99; 多重PE组差异显著, t(20) = 5.02, p < 0.001, d = 1.10, 说明4组被试都成功习得了恐惧, 如图3所示。
图3
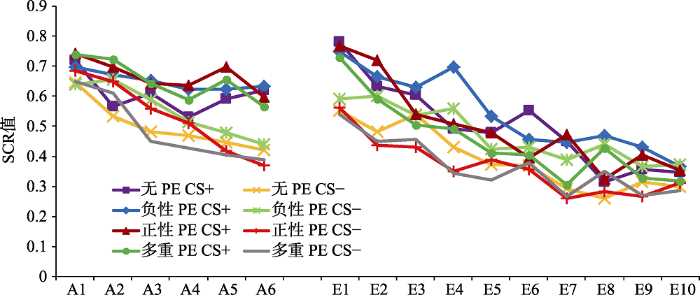
图3
习得、消退阶段各组皮电值的变化
注:横坐标A代表习得阶段(Acquisition), E代表消退阶段(Extinction), 数字代表该阶段的试次。
条件恐惧消退:重复测量方差分析结果显示, 刺激类型的主效应显著, F(1,80) = 49.97, p < 0.001, 偏η2 = 0.38; 试次的主效应显著, F(9,72) = 15.41, p < 0.001, 偏η2 = 0.66; 刺激类型和试次的交互作用不显著, F(9,72) = 1.97, p = 0.06。组别效应不显著, F(3,80) = 0.79, p = 0.50; 刺激类型和组别的交互作用也不显著, F(3,80) = 0.83, p = 0.48, 说明4组被试对CS+和CS-的恐惧反应不存在显著差异。再分别对各组消退阶段最后一个CS+与CS-进行配对样本t检验, 结果显示, 无PE组差异不显著, t(20) = 1.55, p = 0.14; 负性PE组差异不显著, t(20) = -0.10, p = 0.93; 正性PE组差异不显著, t(20) = 0.66, p = 0.51; 多重PE组差异不显著, t(20) = 0.65, p = 0.52, 说明4组被试都进行了成功的消退, 如图3所示。
自发恢复测试:首先对再消退全程进行2(刺激类型) × 2(时间阶段: 前半段, 后半段) × 4(组别)的重复测量方差分析, 结果显示, 刺激类型的主效应显著, F(1,80) = 22.13, p < 0.001, 偏η2 = 0.22; 阶段的主效应显著, F(1,80) = 22.08, p < 0.001, 偏η2 = 0.22; 刺激类型和阶段的交互作用显著, F(1,80) = 11.68, p < 0.01, 偏η2 = 0.13。事后比较发现, 被试只在CS+刺激上出现恐惧的自发恢复。组别效应不显著, F(3,80) = 0.33, p = 0.80; 但刺激类型和组别的交互作用显著, F(3,80) = 3.38, p < 0.05, 偏η2 = 0.11。分离刺激类型单独对CS+进行重复测量方差分析发现, 阶段与组别的交互作用显著, F(3,80) = 3.57, p < 0.05, 偏η2 = 0.13, 说明4组被试对CS+刺激的自发恢复出现差异。
以第三天再消退阶段第一个试次的SCR值和第二天消退阶段最后一个试次的SCR值的变化作为条件性恐惧自发恢复的指标(Schiller et al., 2010)。分别对各组被试两阶段的两个关键试次(第二天消退训练的最后一个试次与第三天再消退的第一个试次)进行配对样本t检验, 结果显示, 无PE组差异显著, t(20) = 3.15, p < 0.05, d = 0.69; 负性PE组差异不显著, t(20) = 0.54, p = 0.59; 正性PE组差异不显著, t(20) = 0.34, p = 0.74; 多重PE组差异显著, t(20) = 2.59, p < 0.05, d = 0.57, 说明只有无PE组和多重PE组出现了恐惧自发恢复, 而负性PE组和正性PE组没有出现恐惧的自发恢复, 如图4所示。为了进一步验证4组被试对CS+刺激恐惧反应的差异, 对各组再消退阶段第一个CS+的SCR值与消退阶段最后一个CS+的SCR值相减, 将两者的差值进行单因素方差分析, 结果显示, 4组被试的SCR差值差异显著, F(3,80) = 3.57, p < 0.05, 多重比较发现, 无PE组和多重PE组的SCR差值显著大于负性PE组和正性PE组, 如图5所示。结果说明, 一个PE组(包括正负性PE)的恐惧自发恢复显著的小于无PE组和多个PE组。
图4
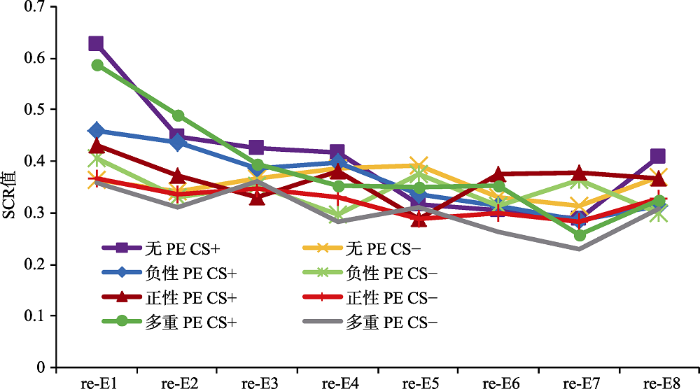
图5
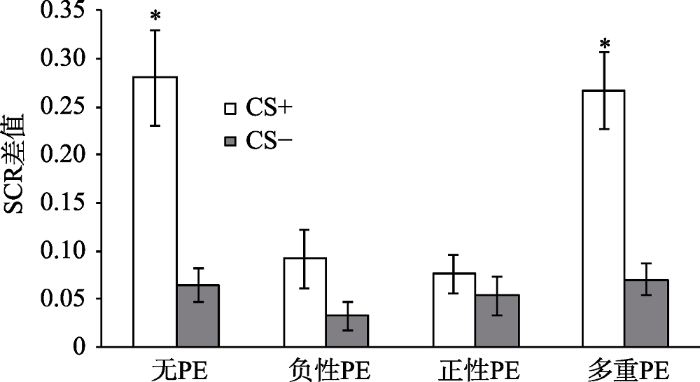
图5
各组自发恢复测试(SCR差值)
注:自发恢复量(SCR差值)=再消退第一个试次SCR值—消退最后一个试次SCR值(Schiller et al., 2010)
恐惧重建测试:首先对最后消退全程进行2(刺激类型) × 2(时间阶段: 前半段, 后半段) × 4(组别)的重复测量方差分析, 结果显示, 刺激类型的主效应显著, F(1,80) = 11.96, p < 0.001, 偏η2 = 0.13; 阶段的主效应显著, F(1,80) = 14.22, p < 0.01, 偏η2 = 0.15; 刺激类型和阶段的交互作用显著, F(1,80) = 4.77, p < 0.05, 偏η2 = 0.06, 事后比较发现, 被试只在CS+刺激上出现恐惧的重建效应。组别效应不显著, F(3,80) = 0.98, p = 0.41; 但刺激类型和组别的交互作用显著, F(3,80) = 3.67, p < 0.05, 偏η2 = 0.12。分离刺激类型单独对CS+进行重复测量方差分析发现, 阶段与组别的交互作用显著, F(3,80) = 3.07, p < 0.05, 偏η2 = 0.10, 说明4组被试对CS+刺激的恐惧重建出现差异。
以第三天重建(单独呈现4个连续的US电击)后的第一个试次的SCR值和第三天再消退阶段最后一个试次的SCR值的变化作为恐惧记忆重建的指标(Schiller et al., 2010)。分别对各组被试两阶段的两个关键试次(重建后消退的第一个试次与再消退的最后一个试次)进行配对样本t检验, 结果显示, 无PE组差异显著, t(20) = 2.85, p < 0.05, d = 0.62; 负性PE组差异不显著, t(20) = -0.46, p = 0.64; 正性PE组差异不显著, t(20) = -0.60, p = 0.56; 多重PE组差异显著, t(20) = 2.12, p < 0.05, d = 0.46, 说明只有无PE组和多重PE组出现了恐惧重建效应, 而负性PE组和正性PE组没有出现恐惧重建效应, 如图6所示。为了进一步验证4组被试对CS+刺激恐惧反应的差异, 对各组重建后第一个CS+的SCR值与再消退阶段最后一个CS+的SCR值相减, 将两者的差值进行单因素方差分析, 结果显示, 4组被试的SCR差值差异显著, F(3,80) = 3.07, p < 0.05, 多重比较发现, 无PE组和多重PE组的SCR差值显著大于负性PE组和正性PE组, 如图7所示。结果说明, 一个PE组(包括正负性PE)的恐惧重建显著的小于无PE组和多个PE组, 变化趋势和恐惧自发恢复指标情况一致。
图6
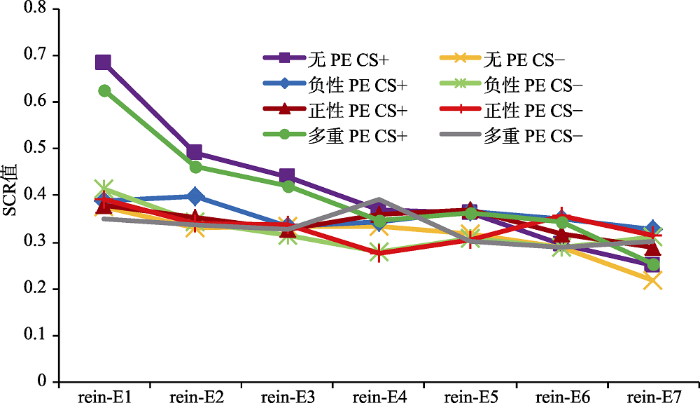
图7
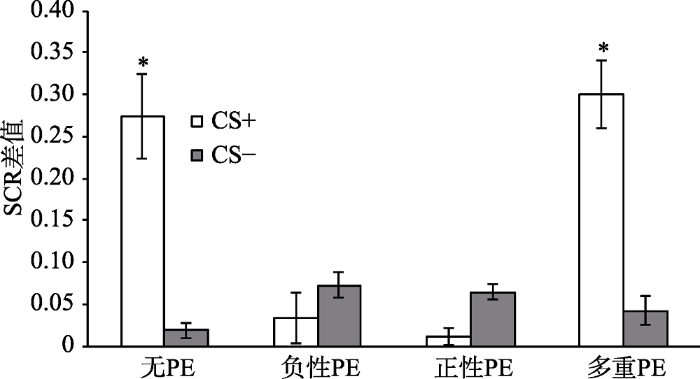
图7
各组重建测试(SCR差值)
注:重建测试(SCR差值)=重建后第一个试次SCR值—再消退最后一个试次SCR值(Schiller et al., 2010)
4 讨论
本研究采用多感官复合刺激(声音+图片)模型作为条件刺激, 借鉴Sevenster等人操纵预期错误的范式(Sevenster et al., 2013, 2014), 通过改变恐惧记忆提取阶段的CS-US配对规则, 形成4种预期错误的情况:无PE、一个负性PE、一个正性PE和多重PE, 比较这4种情况下条件性恐惧记忆提取消退的效果。结果发现, 在4组被试同等程度的习得和消退了恐惧记忆的前提下, 在自发恢复测试中, 无PE组和多重PE组均出现了恐惧记忆的自发恢复, 而负性PE组和正性PE组则没有出现自发恢复。在重建测试中, 无PE组和多重PE组均出现了恐惧记忆的重建效应, 而负性PE组和正性PE组没有出现重建效应; 在恐惧复发的两个指标上效果一致。
4.1 非药物干预预期错误引发的再巩固能够有效阻止复合恐惧记忆返回
目前已有多项研究表明, 记忆提取阶段的预期错误是使记忆进入再巩固的必要条件(Exton- McGuinness et al., 2015; Garrison, Erdeniz, & Done, 2013)。在动物研究中, 采用时间进程性错误(Temporal Error)来产生预期错误, 通过改变非条件刺激在条件刺激中出现的时刻造成预期错误, 也证明了预期错误对于开启记忆再巩固具有关键性作用(Díaz-Mataix et al., 2013), 类似的结果也在不同记忆类型的研究中得到验证(Elsey & Kindt, 2017)。
本研究在前人研究的基础上, 采用跨通道复合刺激记忆模型, 将预期错误应用到提取消退范式中来探究非药物干预记忆再巩固过程是否能够抑制恐惧记忆的返回。实验结果证明利用预期错误对复合刺激进行整体提取后进行行为干预, 依旧能阻止恐惧复发, 该结论验证并拓展了以往研究的结论, 丰富了提取消退范式的理论模型。另外, 本研究采用的是跨通道复合刺激记忆模型, 复合刺激中包含声音刺激与图片刺激, 图片刺激也是多线索模式, 更加贴近现实情境, 这也一定程度上为该范式的临床应用价值提供佐证。
对个体而言, 为了适应外界环境的变化, 记忆必须不断更新(update), 所以再巩固是记忆的自适应过程。由于记忆进行再巩固时必须经历不稳定状态的时间窗, 便于蛋白质重新合成, 在这个不稳定的时间窗内很容易受外界干扰, 因此记忆不会轻易的进入再巩固过程, 除非很有必要更新这个记忆(Beckers & Kindt, 2017)。而当出现预期错误时, 行为的结果不符合原先的预期, 原先的记忆已不具有准确的预测作用, 此时产生新的驱动力促使记忆进行更新。基于记忆再巩固的行为干预模式利用记忆再巩固的动态性特征, 通过各种非侵入性的行为干预制造新信息, 使其与原先记忆进行重新整合更新记忆(Elsey & Kindt, 2017)。行为干预的关键在于必须在再巩固时间窗内实施, 否则没法与原先记忆进行整合。所以, 对于前人研究提取消退范式产生的不一致结果, 或许可以从实验结果中获得一些可能的解释。关于提取消退范式无效的研究中是否是因为没有产生预期错误导致记忆没有进入再巩固过程, 进行的只是传统的消退模式, 所以在测试阶段才会出现恐惧返回, 这个问题值得进一步研究。
4.2 恐惧记忆提取消退的提取边界条件
记忆再巩固的边界条件是指记忆进入再巩固过程中需要满足的条件, 也是保证提取消退等记忆再巩固干预范式有效性的前提。目前大量研究已经证实了一系列边界条件的存在, 如记忆年龄、记忆强度、提取强度、提取次数等(Auber et al., 2013; Nader, 2015; Schwabe et al., 2014)。这些边界条件可以分为两大类, 记忆本身的特性条件和记忆提取阶段的条件。从这一范式的临床应用价值来看, 由于记忆本身的特性例如强度、年龄等往往难以改变, 我们认为研究记忆提取阶段的边界条件, 即对于特定的记忆在提取阶段如何操作才能使其真正进入再巩固, 就显得尤为重要。只有弄清楚提取边界条件是什么以及其作用机制, 我们才能通过实验操作保证记忆能够进入再巩固过程, 进而恐惧记忆才有了改写或清除的可能。所以近年来对于记忆再巩固的边界条件研究越来越多, 其中有观点认为, 先前同类研究中出现了不一致的结果, 是因为它们在提取时新呈现的信息与先前编码的信息的区别程度是不一样的, 即存在信息差异性(Beckers & Kindt, 2017; Elsey & Kindt, 2017)。信息差异是指记忆被再次激活时是需要有新信息存在的, 也就是说, 在提取过程中如果没有出现信息差异, 便不能有效的开启再巩固过程。如果从提取阶段的信息差异这个角度整合再巩固边界条件, 那么对于预期错误的研究就是一个很好的切入点。以往研究多是在类别上界定边界条件, 很少从量上去探讨, 但是要想明确再巩固的边界条件, 对于量的探讨是很有意义的。本研究结果表明, 当提取阶段预期错误的量过大时(实验中多重PE组), 恐惧记忆在两个指标上都有显著的复发, 与无PE组类似, 这说明原始记忆并没有经历再巩固过程, 即提取失败。这一结果与Sevenster等人(2014)的研究结果一致, 再次说明了需要对于预期错误等记忆再巩固的边界条件进行量化的探索(Sevenster et al., 2014)。这无疑增加了临床应用的复杂性, 也是研究中需要解决的难点问题。
4.3 正性、负性预期错误在开启记忆再巩固上的作用
实验结果表明, 单个正性PE和负性PE都可以开启再巩固过程, 虽然预期错误的正负性在提取阶段设置时会存在操作差异, 负性预期错误组提取时呈现两个不带电击的CS+, 而正性预期错误提取是呈现两个带电击的CS+, 但在行为层面上具有同样的效果。有研究者认为, 负性PE和正性PE只是产生预期错误的方式不同, 其结果都是产生了预期错误开启记忆再巩固, 这两种预期错误并没有本质上的差异(Sevenster et al., 2013)。但Spoormaker 等人(2011)利用磁共振脑成像技术(MRI)研究正性预期错误和负性预期错误的差异时发现, 负性PE信号在大脑产生的神经功能连接与正性PE信号产生的神经连接有差异(Spoormaker et al., 2011)。Hon, Das和Kamboj (2016)的研究中将预期错误分为改变效价的预期错误(value-prediction error)和直接省略的预期错误(omission-prediction error), 两类预期错误类型却出现了不同的结果, 说明了预期错误开启记忆再巩固具有类型特异性(Hon et al., 2016)。本研究整合了正性预期错误和负性预期错误两种类型, 在恐惧返回的行为指标上没有发现差异, 结合前人的研究我们推测二者可能只存在神经过程上的差异。因此进一步弄清正性、负性PE提取记忆的作用机制, 还需要大量神经生理机制方面的研究。
4.4 适当的预期错误才能开启记忆再巩固
提取阶段的预期错误引发记忆再巩固也存在一定的限制, 预期错误过大时并不能够使记忆进入再巩固, 不能成功提取记忆, 从而导致恐惧返回。实验中多重PE组在提取时出现了两个预期错误, 结果第三天出现了明显的自发恢复和重建, 说明记忆没有进入再巩固过程, 只是进行了传统消退。实验结果与Sevenster等人(2014)的研究结果一致, 研究者认为这可能是因为多次出现和预期不一样的结果, 个体会认为新信息和原来的信息没有关系, 没有激活原始记忆, 而是会习得新的记忆形成两类记忆(Sevenster et al., 2014)。Li等人(2017)在研究复合CS提取比例对提取消退效果的影响中发现, 使用中等比例的CS的提取效果是最好的, 全比例提取不能产生预期错误, 小比例提取则会产生过小的预期错误, 这也从侧面说明存在一个最佳的预期错误量(Li et al., 2017)。所以, 提取时出现预期错误的量可以界定记忆是选择再巩固更新记忆还是重新形成新的记忆。Elsey和Kindt (2017)指出, 当对记忆进行提取之后, 记忆会有几种可能的变化方向, 包括:仅提取, 形成消退记忆, 激活进入不稳定状态等。而只有产生合适的预期错误才能使记忆激活进入不稳定, 为记忆更新提供条件(Elsey & Kindt, 2017)。在临床治疗中, 如果提取时能够使患者产生适当预期错误将会增强治疗效果(Beckers & Kindt, 2017)。但目前对于这部分的研究很有限, 如何引发和维持适当的预期错误也是临床应用面临的一个难点。未来还需大量的基础研究对预期错误的量化效应进行更深入的探索。
4.5 总结与展望
本研究在前人研究的基础上, 以复合条件性恐惧记忆为模型, 以人为被试, 将预期错误应用在提取消退范式中, 证明了提取时出现单个预期错误能够开启记忆再巩固从而充分发挥提取消退范式消退恐惧记忆抑制恐惧返回的效果, 如果提取时没有出现预期错误或者出现了过量预期错误则提取消退范式没有起到抑制恐惧返回的效果。但本研究尚不明确多大范围程度上的预期错误能够开启记忆再巩固。对于预期错误开启记忆再巩固, 是否存在一个量的区间, 只要在这个区间内设置的预期错误都能成功开启再巩固, 而且这个区间是否会因为记忆类型等外界条件而改变, 这一现象出现的神经机制是怎样的等等, 都需要未来进一步的研究。
因此, 未来应在基础研究中探索记忆再巩固边界条件更细化的量化分类, 甚至是具体的预测指标, 来判断记忆是否激活进入了再巩固, 进而探索如何在临床治疗环境中让被试产生适当的预期错误。这是研究上的一个难点, 也是通往临床治疗的过程中需要进一步解决的关键问题。
参考文献
Post-retrieval extinction as reconsolidation interference: Methodological issues or boundary conditions?
DOI:10.1007/s00213-013-3004-1
URL
PMID:23404065
[本文引用: 2]

Memories that are emotionally arousing generally promote the survival of species; however, the systems that modulate emotional learning can go awry, resulting in pathological conditions such as post-traumatic stress disorders, phobias, and addiction. Understanding the conditions under which emotional memories can be targeted is a major research focus as the potential to translate these methods into clinical populations carries important implications. It has been demonstrated that both fear and drug-related memories can be destabilised at their retrieval and require reconsolidation to be maintained. Therefore, memory reconsolidation offers a potential target period during which the aberrant memories underlying psychiatric disorders can be disrupted. Monfils et al. (Science 324:951 955, 2009 ) have shown for the first time that safe information provided through an extinction session after retrieval (during the reconsolidation window) may update the original memory trace and prevent the return of fear in rats. In recent years, several authors have then tested the effect of post-retrieval extinction on reconsolidation of either fear or drug-related memories in both laboratory animals and humans. In this article, we review the literature on post-reactivation extinction, discuss the differences across studies on the methodological ground, and review the potential boundary conditions that may explain existing discrepancies and limit the potential application of post-reactivation extinction approaches.
Memory reconsolidation interference as an emerging treatment for emotional disorders: Strengths, limitations, challenges, and opportunities
DOI:10.1146/annurev-clinpsy-032816-045209
URL
PMID:5424072
[本文引用: 4]
Experimental research on emotional memory reconsolidation interference, or the induction of amnesia for previously established emotional memory, has a long tradition, but the potential of that research for the development of novel interventions to treat psychological disorders has been recognized only recently. Here we provide an overview of basic research and clinical studies on emotional memory reconsolidation interference. We point out specific advantages of interventions based on memory reconsolidation interference over traditional treatment for emotional disorders. We also explain how findings from basic research suggest limitations and challenges to clinical translation that may help to understand why clinical trials have met with mixed success so far and how their success can be increased. In closing, we preview new intervention approaches beyond the induction of amnesia that the phenomenon of memory reconsolidation may afford for alleviating the burden imposed by emotional memories and comment on theoretical controversies regarding the nature of memory reconsolidation.
Sources of relapse after extinction in Pavlovian and instrumental learning
DOI:10.1016/0272-7358(91)90091-8
URL
[本文引用: 1]
Extinction may be related to therapeutic procedures designed to eliminate unwanted thoughts, emotions, or behaviors. This article reviews animal learning research which suggests that extinction does not erase the original learning, but rather makes behavior especially sensitive to the background, or context, in which it occurs. Context-dependence of responding is evident after extinction in both Pavlovian and instrumental (operant) learning. In either case, memories of conditioning and extinction are both retained through extinction, and are available for retrieval by appropriate retrieval cues. When the context promotes retrieval of extinction, extinction performance is observed. But when the context is changed, or when the current context promotes retrieval of conditioning, conditioning performance is often restored. These effects are produced by contexts provided by stimuli as diverse as physical environments, reinforcer after-effects, drug states, emotions, and the passage of time. The results have implications for relapse, and its prevention, following therapy.
Treatment of specific phobia in adults
DOI:10.1016/j.cpr.2006.10.002
URL
PMID:17112646
[本文引用: 1]
This is a comprehensive review of treatment studies in specific phobia. Acute and long-term efficacy studies of -clycoserine. Despite the acute benefits of in vivo exposure, greater attention should be paid to improve treatment acceptance and retention, and additional controlled studies of more acceptable treatments are needed.
Role of the amygdala in fear extinction measured with potentiated startle
DOI:10.1111/j.1749-6632.2003.tb07084.x
URL
PMID:12724161
[本文引用: 1]
Abstract A bstract : Although much is now known about the neural basis of excitatory fear conditioning, much less is known about the neural basis of inhibitory conditioning. One type of inhibitory conditioning is extinction, a process in which stimuli that elicit fear by virtue of previous associations with aversive stimuli such as shock (excitatory fear conditioning) are now presented in the absence of the aversive stimuli (extinction training). As a result, the ability of the conditioned stimulus to elicit fear gradually diminishes. Extinction is different from forgetting and does not reflect an erasure of the original fear memory. Instead, extinction is an active form of inhibitory learning that competes with excitatory fear conditioning. Infusions into the amygdala (a brain area essential for excitatory fear conditioning) of either NMDA receptor antagonists or inhibitors of the NMDA-receptor-linked mitogen-activated protein kinase cascade block extinction learning. Conversely, the NMDA receptor agonist D-cycloserine facilitates extinction after either systemic administration or intra-amygdala infusion. Because therapeutic interventions based on extinction procedures are commonly used to treat fear disorders, and because D-cycloserine is a widely available and safe compound, D-cycloserine or similar agents might be usefully combined with traditional extinction-based approaches in the treatment of clinical fear.
Detection of a temporal error triggers reconsolidation of amygdala-dependent memories
DOI:10.1016/j.cub.2013.01.053
URL
PMID:23453952
[本文引用: 2]
78 Fear reconsolidation is only triggered when memory is updated with new information 78 Time is a core part of the Pavlovian CS-US association 78 Reactivation synaptic plasticity occurs only when error prediction is detected
Tackling maladaptive memories through reconsolidation: From neural to clinical science
DOI:10.1016/j.nlm.2017.03.007
URL
[本文引用: 5]
Behavioral neuroscience has greatly informed how we understand the formation, persistence, and plasticity of memory. Research has demonstrated that memory reactivation can induce a labile period, during which previously consolidated memories are sensitive to change, and in need of restabilization. This process is known as reconsolidation. Such findings have advanced not only our basic understanding of memory processes, but also hint at the prospect of harnessing these insights for the development of a new generation of treatments for disorders of emotional memory. However, even in simple experimental models, the conditions for inducing memory reconsolidation are complex: memory labilization appears to result from the interplay of learning history, reactivation, and also individual differences, posing difficulties for the translation of basic experimental research into effective clinical interventions. In this paper, we review a selection of influential animal and human research on memory reconsolidation to illustrate key insights these studies afford. We then consider how these findings can inform the development of new treatment approaches, with a particular focus on the transition of memory from reactivation, to reconsolidation, to new memory formation, as well as highlighting possible limitations of experimental models. If the challenges of translational research can be overcome, and if reconsolidation-based procedures become a viable treatment option, then they would be one of the first mental health treatments to be directly derived from basic neuroscience research. This would surely be a triumph for the scientific study of mind and brain.
Updating memories--the role of prediction errors in memory reconsolidation
DOI:10.1016/j.bbr.2014.10.011
URL
PMID:25453746
[本文引用: 2]
Memories are not static imprints of past experience, but rather are dynamic entities which enable us to predict outcomes of future situations and inform appropriate behaviours. In order to maintain the relevance of existing memories to our daily lives, memories can be updated with new information via a process of reconsolidation. In this review we describe recent experimental advances in the reconsolidation of both appetitive and aversive memory, and explore the neuronal mechanisms that underpin the conditions under which reconsolidation will occur. We propose that a prediction error signal, originating from dopaminergic midbrain neurons, is necessary for destabilisation and subsequent reconsolidation of a memory.
The fate of memory: Reconsolidation and the case of Prediction Error
DOI:10.1016/j.neubiorev.2016.06.004
URL
PMID:27287939
[本文引用: 1]
The ability to make predictions based on stored information is a general coding strategy. A Prediction-Error (PE) is a mismatch between expected and current events. It was proposed as the process by which memories are acquired. But, our memories like ourselves are subject to change. Thus, an acquired memory can become active and update its content or strength by a labilization-reconsolidation process. Within the reconsolidation framework, PE drives the updating of consolidated memories. Moreover, memory features, such as strength and age, are crucial boundary conditions that limit the initiation of the reconsolidation process. In order to disentangle these boundary conditions, we review the role of surprise, classical models of conditioning, and their neural correlates. Several forms of PE were found to be capable of inducing memory labilization-reconsolidation. Notably, many of the PE findings mirror those of memory-reconsolidation, suggesting a strong link between these signals and memory process. Altogether, the aim of the present work is to integrate a psychological and neuroscientific analysis of PE into a general framework for memory-reconsolidation.
Prediction error in reinforcement learning: A meta-analysis of neuroimaging studies
DOI:10.1016/j.neubiorev.2013.03.023
URL
PMID:23567522
[本文引用: 1]
Activation likelihood estimation (ALE) meta-analyses were used to examine the neural correlates of prediction error in reinforcement learning. The findings are interpreted in the light of current computational models of learning and action selection. In this context, particular consideration is given to the comparison of activation patterns from studies using instrumental and Pavlovian conditioning, and where reinforcement involved rewarding or punishing feedback. The striatum was the key brain area encoding for prediction error, with activity encompassing dorsal and ventral regions for instrumental and Pavlovian reinforcement alike, a finding which challenges the functional separation of the striatum into a dorsal 'actor' and a ventral 'critic'. Prediction error activity was further observed in diverse areas of predominantly anterior cerebral cortex including medial prefrontal cortex and anterior cingulate cortex. Distinct patterns of prediction error activity were found for studies using rewarding and aversive reinforcers; reward prediction errors were observed primarily in the striatum while aversive prediction errors were found more widely including insula and habenula.
The effects of cognitive reappraisal following retrieval-procedures designed to destabilize alcohol memories in high-risk drinkers
DOI:10.1007/s00213-015-4164-y
URL
PMID:4751186
[本文引用: 1]
Rationale Addiction is a disorder of motivational learning and memory. Maladaptive motivational memories linking drug-associated stimuli to drug seeking are formed over hundreds of reinf
An isolated retrieval trial before extinction session does not prevent the return of fear
DOI:10.1016/j.bbr.2015.03.052
URL
PMID:25827926
[本文引用: 1]
Several studies have shown that an isolated retrieval trial before the extinction session (retrieval-extinction) prevents the return of fear memory by inhibition of reconsolidation. Other studies have reported that retrieval-extinction did not prevent the return of the fear. To date, it is still unclear whether retrieval-extinction prevents the return of the original fear memory. A previous study revealed that reconsolidation of conditioned fear memory was not induced by the brevity of the retrieval session. Thus, we examined whether the number of retrievals in the retrieval-extinction paradigm was involved in the prevention of return of fear (Experiment 1). Furthermore, studies with different-age experimental subjects have shown conflicting results. We investigated the potential impact of age on the inhibitory effect of retrieval-extinction on the return of fear (Experiment 2). Our major findings were as follows: (1) Retrieval-extinction procedure did not prevent the return of fear, regardless of the intensity (number of presentations) of the stimulus inducing retrieval of fear memory. (2) The mice in both juvenile and adult age groups (4 and 8 weeks old) retrieved fear memory after retrieval-extinction. These results suggest the possibility that extinction after retrieval does not inhibit reconsolidation of previously consolidated fear memory.
Reconsolidation in a human fear conditioning study: A test of extinction as updating mechanism
DOI:10.1016/j.biopsycho.2011.09.016
URL
PMID:21986472
[本文引用: 1]
Abstract Disrupting reconsolidation seems to be a promising approach to dampen the expression of fear memory. Recently, we demonstrated that disrupting reconsolidation by a pharmacological manipulation specifically targeted the emotional expression of memory (i.e., startle response). Here we test in a human differential fear-conditioning paradigm with fear-relevant stimuli whether the spacing of a single unreinforced retrieval trial relative to extinction learning allows for "rewriting" the original fear association, thereby preventing the return of fear. In contrast to previous findings reported by Schiller et al. (2010), who used a single-method for indexing fear (skin conductance response) and fear-irrelevant stimuli, we found that extinction learning within the reconsolidation window did not prevent the recovery of fear on multiple indices of conditioned responding (startle response, skin conductance response and US-expectancy). These conflicting results ask for further critical testing given the potential impact on the field of emotional memory and its application to clinical practice. Copyright 2011 Elsevier B.V. All rights reserved.
Independent cellular processes for hippocampal memory consolidation and reconsolidation
DOI:10.1126/science.1095760
URL
PMID:15073322
[本文引用: 1]
The idea that new memories undergo a time-dependent consolidation process after acquisition has received considerable experimental support. More controversial has been the demonstration that established memories, once recalled, become labile and sensitive to disruption, requiring "reconsolidation" to become permanent. By infusing antisense oligodeoxynucleotides into the hippocampus of rats, we show that consolidation and reconsolidation are doubly dissociable component processes of memory. Consolidation involves brain-derived neurotrophic factor (BDNF) but not the transcription factor Zif268, whereas reconsolidation recruits Zif268 but not BDNF. These findings confirm a requirement for BDNF specifically in memory consolidation and also resolve the role of Zif268 in brain plasticity, learning, and memory.
An update on memory reconsolidation updating
DOI:10.1016/j.tics.2017.04.006
URL
PMID:28495311
[本文引用: 1]
The reactivation of a stored memory in the brain can make the memory transiently labile. During the time it takes for the memory to restabilize (reconsolidate) the memory can either be reduced by an amnesic agent or enhanced by memory enhancers. The change in memory expression is related to changes in the brain correlates of long-term memory. Many have suggested that such retrieval-induced plasticity is ideally placed to enable memories to be updated with new information. This hypothesis has been tested experimentally, with a translational perspective, by attempts to update maladaptive memories to reduce their problematic impact. We review here progress on reconsolidation updating studies, highlighting their translational exploitation and addressing recent challenges to the reconsolidation field.
Moderate partially reduplicated conditioned stimuli as retrieval cue can increase effect on preventing relapse of fear to compound stimuli
DOI:10.3389/fnhum.2017.00575
URL
PMID:5714856
[本文引用: 2]
Abstract The theory of memory reconsolidation argues that consolidated memory is not unchangeable. Once a memory is reactivated it may go back into an unstable state and need new protein synthesis to be consolidated again, which is called "memory reconsolidation". Boundary studies have shown that interfering with reconsolidation through pharmacologic or behavioral intervention can lead to the updating of the initial memory, for example, erasing undesired memories. Behavioral procedures based on memory reconsolidation interference have been shown to be an effective way to inhibit fear memory relapse after extinction. However, the effectiveness of retrieval-extinction differs by subtle differences in the protocol of the reactivation session. This represents a challenge with regard to finding an optimal operational model to facilitate its clinical use for patients suffering from pathogenic memories such as those associated with post-traumatic stress disorder. Most of the laboratory models for fear learning have used a single conditioned stimulus (CS) paired with an unconditioned stimulus (US). This has simplified the real situation of traumatic events to an excessive degree, and thus, limits the clinical application of the findings based on these models. Here, we used a basic visual compound CS model as the CS to ascertain whether partial repetition of the compound CSs in conditioning can reactivate memory into reconsolidation. The results showed that the no retrieval group or the 1/3 ratio retrieval group failed to open the memory reconsolidation time window. The 2/3 repetition retrieval group and the whole repetition retrieval group were able to prevent fear reinstatement, whereas only a 2/3 ratio repetition of the initial compound CS as a reminder could inhibit spontaneous recovery. We inferred that a retrieval-extinction paradigm was also effective in a more complex model of fear if a sufficient prediction error (PE) could be generated in the reactivation period. In addition, in order to achieve an optimal effect, a CS of moderate discrepancy should be used as a reminder.
The conditions that promote fear learning: Prediction error and Pavlovian fear conditioning
DOI:10.1016/j.nlm.2013.05.002
URL
PMID:23684989
[本文引用: 1]
A key insight of associative learning theory is that learning depends on the actions of prediction error: a discrepancy between the actual and expected outcomes of a conditioning trial. When positive, such error causes increments in associative strength and, when negative, such error causes decrements in associative strength. Prediction error can act directly on fear learning by determining the effectiveness of the aversive unconditioned stimulus or indirectly by determining the effectiveness, or associability, of the conditioned stimulus. Evidence from a variety of experimental preparations in human and non-human animals suggest that discrete neural circuits code for these actions of prediction error during fear learning. Here we review the circuits and brain regions contributing to the neural coding of prediction error during fear learning and highlight areas of research (safety learning, extinction, and reconsolidation) that may profit from this approach to understanding learning.
The establishment and assessment of a native affective sound system
本土化情绪声音库的编制和评定
DOI:10.3969/j.issn.1671-6981.2006.02.040
URL
[本文引用: 1]
为情绪研究提供一套本土量化的声音刺激材料,在大量收集各种声音刺激的基础上精选出453个声音,并征集50名大学生对声音的愉悦度、唤醒度和优势度进行了自我报告的9点量表评分。结果表明男女生对部分声音的情绪感受有所不同;聚类分析可将声音聚为6类,大致可引发愉快、悲伤、恐惧、厌恶等情绪,提示可以同时从情绪维度和情绪类型两种途径研究情绪问题,两者结果也有相互比较的可能性;本情绪声音库还有待进一步完善。
An unconditioned stimulus retrieval extinction procedure to prevent the return of fear memory
DOI:10.1016/j.biopsych.2014.03.027
URL
PMID:24813334
[本文引用: 1]
Conditioned fear memories can be updated by extinction during reconsolidation, and this effect is specific to the reactivated conditioned stimulus (CS). However, a traumatic event can be associated with several cues, and each cue can potentially trigger recollection of the event. We introduced a technique to target all diverse cues associated with an aversive event that causes fear.In human experiments, 161 subjects underwent modified fear conditioning, in which they were exposed to an unconditioned stimulus (US) or unreinforced CS to reactivate the memory and then underwent extinction, spontaneous recovery, and reinstatement. In animal experiments, 343 rats underwent contextual fear conditioning under a similar protocol as that used in the human experiments. We also explored the molecular alterations after US reactivation in rats.Presentation of a lower intensity US before extinction disrupted the associations between the different CS and reactivated US in both humans and rats. This effect persisted for at least 6 months in humans and was selective to the reactivated US. This procedure was also effective for remote memories in both humans and rats. Compared with the CS, the US induced stronger endocytosis of alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid glutamate receptors 1 and 2 and stronger activation of protein kinase A, p70S6 kinase, and cyclic adenosine monophosphate response element binding protein in the dorsal hippocampus in rats.These findings demonstrate that a modified US retrieval extinction strategy may have a potential impact on therapeutic approaches to prevent the return of fear.
Don’t fear ‘fear conditioning’: Methodological considerations for the design and analysis of studies on human fear acquisition, extinction, and return of fear
DOI:10.1016/j.neubiorev.2017.02.026
URL
PMID:28263758
[本文引用: 1]
Abstract The so-called 'replicability crisis' has sparked methodological discussions in many areas of science in general, and in psychology in particular. This has led to recent endeavours to promote the transparency, rigour, and ultimately, replicability of research. Originating from this zeitgeist, the challenge to discuss critical issues on terminology, design, methods, and analysis considerations in fear conditioning research is taken up by this work, which involved representatives from fourteen of the major human fear conditioning laboratories in Europe. This compendium is intended to provide a basis for the development of a common procedural and terminology framework for the field of human fear conditioning. Whenever possible, we give general recommendations. When this is not feasible, we provide evidence-based guidance for methodological decisions on study design, outcome measures, and analyses. Importantly, this work is also intended to raise awareness and initiate discussions on crucial questions with respect to data collection, processing, statistical analyses, the impact of subtle procedural changes, and data reporting specifically tailored to the research on fear conditioning.
A novel UCS memory retrieval-extinction procedure to inhibit relapse to drug seeking
DOI:10.1038/ncomms8675
URL
PMID:4510700
[本文引用: 1]
We recently reported that a conditioned stimulus (CS) memory retrieval-extinction procedure decreases reinstatement of cocaine and heroin seeking in rats and heroin craving in humans. Here we show that non-contingent cocaine or methylphenidate injections (UCS retrieval) 1 before the extinction sessions decreases cocaine-priming-induced reinstatement, spontaneous recovery, and renewal of cocaine seeking in rats. Unlike the CS-based memory retrieval-extinction procedure, the UCS memory retrieval manipulation decreases renewal and reinstatement of cocaine seeking in the presence of cocaine cues that were not present during extinction training and also decreases cocaine seeking when the procedure commences after 28 days of abstinence. The inhibitory effect of the UCS retrieval manipulation on cocaine-priming-induced reinstatement is mediated by regulation of AMPA-receptor endocytosis in the basolateral amygdala. The UCS memory retrieval-extinction procedure has superior relapse prevention characteristics than the CS memory retrieval-extinction procedure and could be a promising method for decreasing relapse in human addicts. Cue-based therapies for treating drug addiction have proven to be only partially effective. Here the authors demonstrate a new memory retrieval based treatment protocol for drug addiction that results in long-lasting inhibition of drug seeking behavior in rodents.
Memory-a century of consolidation
DOI:10.1126/science.287.5451.248 URL [本文引用: 1]
Extinction-reconsolidation boundaries: Key to persistent attenuation of fear memories
DOI:10.1126/science.1167975
URL
PMID:3625935
[本文引用: 2]
Dysregulation of the fear system is at the core of many psychiatric disorders. Much progress has been made in uncovering the neural basis of fear learning through studies in which associative emotional memories are formed by pairing an initially neutral stimulus (conditioned stimulus, CS; e.g., a tone) to an unconditioned stimulus (US; e.g., a shock). Despite recent advances, the question of how to persistently weaken aversive CS-US associations, or dampen traumatic memories in pathological cases, remains a major dilemma. Two paradigms (blockade of reconsolidation and extinction) have been used in the laboratory to reduce acquired fear. Unfortunately, their clinical efficacy is limited: Reconsolidation blockade typically requires potentially toxic drugs, and extinction is not permanent. Here, we describe a behavioral design in which a fear memory in rats is destabilized and reinterpreted as safe by presenting an isolated retrieval trial before an extinction session. This procedure permanently attenuates the fear memory without the use of drugs.
Mechanisms of fear extinction
DOI:10.1038/sj.mp.4001939 URL [本文引用: 1]
Reconsolidation and the dynamic nature of memory
DOI:10.1007/978-3-319-24364-1_1
URL
PMID:26354895
[本文引用: 2]
Research on the reconsolidation effect was greatly revitalized by the highly analytic demonstration of memory reconsolidation (Nader et al. 2000) in a well-defined behavioral protocol (auditory fear...
The labile nature of consolidation theory
DOI:10.1038/35044580
URL
PMID:11257912
[本文引用: 1]
'Consolidation' has been used to describe distinct but related processes. In considering the implications of our recent findings on the lability of reactivated fear memories, we view consolidation and reconsolidation in terms of molecular events taking place within neurons as opposed to interactions between brain regions. Our findings open up a new dimension in the study of memory consolidation. We argue that consolidation is not a one-time event, but instead is reiterated with subsequent activation of the memories.
A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement
In A. H. Black & W. F. Prokasy (Eds.),Publication » A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement.
Preventing the return of fear in humans using reconsolidation update mechanisms
DOI:10.1038/nature08637
URL
PMID:3640262
[本文引用: 7]
Recent research on changing fears has examined targeting reconsolidation. During reconsolidation, stored information is rendered labile after being retrieved. Pharmacological manipulations at this stage result in an inability to retrieve the memories at later times, suggesting that they are erased or persistently inhibited. Unfortunately, the use of these pharmacological manipulations in humans can be problematic. Here we introduce a non-invasive technique to target the reconsolidation of fear memories in humans. We provide evidence that old fear memories can be updated with non-fearful information provided during the reconsolidation window. As a consequence, fear responses are no longer expressed, an effect that lasted at least a year and was selective only to reactivated memories without affecting others. These findings demonstrate the adaptive role of reconsolidation as a window of opportunity to rewrite emotional memories, and suggest a non-invasive technique that can be used safely in humans to prevent the return of fear.
Reconsolidation of human memory: Brain mechanisms and clinical relevance
DOI:10.1016/j.biopsych.2014.03.008
URL
PMID:24755493
[本文引用: 2]
The processes of memory formation and storage are complex and highly dynamic. Once memories are consolidated, they are not necessarily fixed but can be changed long after storage. In particular, seemingly stable memories may re-enter an unstable state when they are retrieved, from which they must be re-stabilized during a process known as reconsolidation. During reconsolidation, memories are susceptible to modifications again, thus providing an opportunity to update seemingly stable memories. While initial demonstrations of memory reconsolidation came mainly from animal studies, evidence for reconsolidation in humans is now accumulating as well. Here, we review recent advances in our understanding of human memory reconsolidation. After a summary of findings on the reconsolidation of human fear and episodic memory, we focus particularly on recent neuroimaging data that provide first insights into how reconsolidation processes are implemented in the human brain. Finally, we discuss the implications of memory modifications during reconsolidation for the treatment of mental disorders such as posttraumatic stress disorder and drug addiction.
Prediction error governs pharmacologically induced amnesia for learned fear
DOI:10.1126/science.1231357 URL [本文引用: 5]
Prediction error demarcates the transition from retrieval, to reconsolidation, to new learning
DOI:10.1101/lm.035493.114
URL
PMID:25320349
[本文引用: 7]
Although disrupting reconsolidation is promising in targeting emotional memories, the conditions under which memory becomes labile are still unclear. The current study showed that post-retrieval changes in expectancy as an index for prediction error may serve as a read-out for the underlying processes engaged by memory reactivation. Minor environmental changes define whether retrieval induces memory reconsolidation or the initiation of a new memory trace even before fear extinction can be observed.
Disrupting reconsolidation: Pharmacological and behavioral manipulations
DOI:10.1101/lm.2148511
URL
PMID:21576515
[本文引用: 1]
We previously demonstrated that disrupting reconsolidation by pharmacological manipulations "deleted" the emotional expression of a fear memory in humans. If we are to target reconsolidation in patients with anxiety disorders, the disruption of reconsolidation should produce content-limited modifications. At the same time, the fear-erasing effects should not be restricted to the feared cue itself considering that fear generalization is a main characteristic of anxiety disorders. In Experiment I and Experiment I(b), we addressed these issues using a within-subject differential startle fear conditioning paradigm and a test of fear generalization. In Experiment II, we tested whether a behavioral approach targeting the reconsolidation through extinction learning was also effective in weakening the original fear memory. A behavioral procedure is evidently preferred over drug manipulations provided that similar effects can be obtained. Here, the extinction procedure subsequent to retrieval did not "erase" the emotional expression of the fear memory as the retrieval techniques (i.e., reminder shocks and reacquisition) unveiled a return of the startle fear response to the fear-relevant stimuli. In contrast, -adrenergic receptor blockade during reconsolidation selectively deleted the fear-arousing aspects of the memory (i.e., startle fear response) along with its category-related information. The pharmacological manipulation rendered the core memory trace too weak to observe fear generalization after successful reacquisition. Hence, relearning following the disruption of reconsolidation seems to be qualitatively different from initial learning. Our findings demonstrate that disrupting reconsolidation by pharmacological manipulations, although selective, undermines the generalization of fear, a key feature of anxiety disorders.
The neural correlates of negative prediction error signaling in human fear conditioning
DOI:10.1016/j.neuroimage.2010.09.042
URL
PMID:20869454
[本文引用: 1]
In a temporal difference (TD) learning approach to classical conditioning, a prediction error (PE) signal shifts from outcome deliverance to the onset of the conditioned stimulus. Omission of an expected outcome results in a negative PE signal, which is the initial step towards successful extinction. In order to visualize negative PE signaling during fear conditioning, we employed combined functional magnetic resonance (fMRI) and skin conductance response (SCR) measurements in a conditioning task with visual stimuli and mild electrical shocks. Positive PE signaling was associated with increased activation in the bilateral insula, supplementary motor area, brainstem, and visual cortices. Negative PE signaling was associated with increased activation in the ventromedial and dorsolateral prefrontal cortices, the left lateral orbital gyrus, the middle temporal gyri, angular gyri, and visual cortices. The involvement of the ventromedial prefrontal and orbitofrontal cortex in extinction learning has been well documented, and this study provides evidence for the notion that these regions are already involved in negative PE signaling during fear conditioning.Research Highlights? The orbitofrontal and ventromedial prefrontal cortex signal shock omission in fear conditioning. ? Different temporal difference models reveal similar statistical maps of activation. ? The stimulus-offset skin conductance response may be a sensitive measure of fear conditioning. ? The neural circuitry of fear extinction is involved in prediction error signaling during conditioning.
A memory retrieval-extinction procedure to prevent drug craving and relapse
DOI:10.1126/science.1215070 URL [本文引用: 1]
The effect of compound stimulus to conditioned fear extinction in retrieval-extinction paradigm
提取-消退范式中复合刺激对恐惧消退的影响

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}